

Photoreceptor responses of fruitflies with normal and reduced arrestin content studied by simultaneous measurements of visual pigment fluorescence and ERG
- PMID: 19924417
- PMCID: PMC2797847
- DOI: 10.1007/s00359-009-0489-5
Photoreceptor responses of fruitflies with normal and reduced arrestin content studied by simultaneous measurements of visual pigment fluorescence and ERG
Abstract
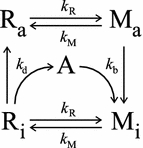
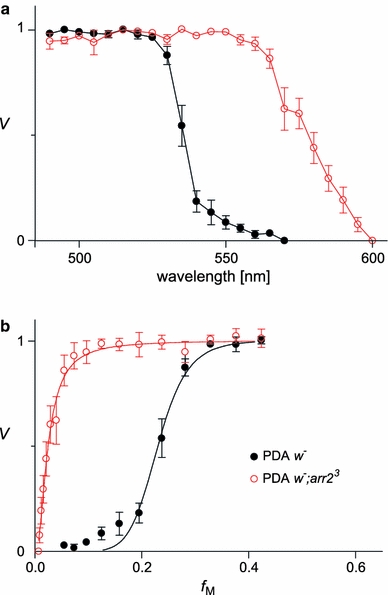
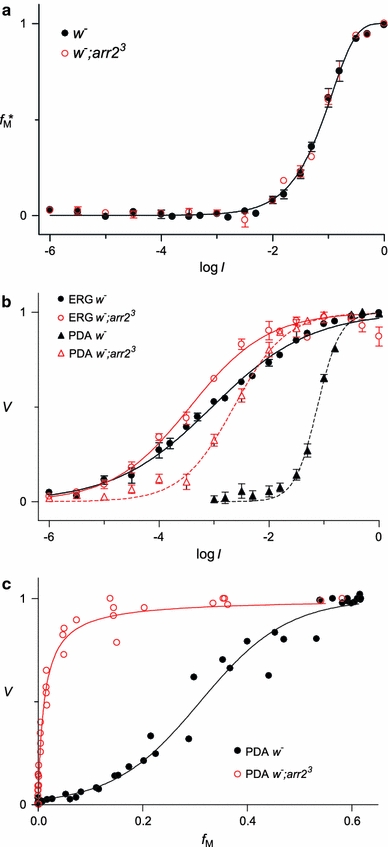
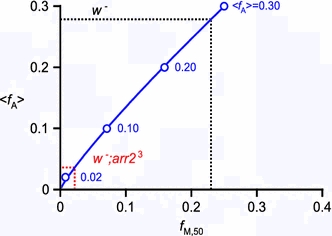
We have simultaneously measured the electroretinogram (ERG) and the metarhodopsin content via fluorescence in white-eyed, wild-type Drosophila and the arrestin2 hypomorphic mutant (w(-);arr2 (3)) at a range of stimulus wavelengths and intensities. Photoreceptor response amplitude and termination (transition between full repolarization and prolonged depolarizing afterpotential, PDA) were related to visual pigment conversions and arrestin concentration. The data were implemented in a kinetic model of the rhodopsin-arrestin cycle, allowing us to estimate the active metarhodopsin concentration as a function of effective light intensity and arrestin concentration. Arrestin reduction in the mutant modestly increased the light sensitivity and decreased the photoreceptor dynamic range. Compared to the wild type, in the mutant the transition between full repolarization and PDA occurred at a lower metarhodopsin fraction and was more abrupt. We developed a steady-state stochastic model to interpret the dependence of the PDA on effective light intensity and arrestin content and to help deduce the arrestin to rhodopsin ratio from the sensitivity and PDA data. The feasibility of different experimental methods for the estimation of arrestin content from ERG and PDA is discussed.
Figures





References
-
- Cosens DJ, Briscoe D. A switch phenomenon in the compound eye of the white-eyed mutant of Drosophila melanogaster. J Insect Physiol. 1972;18:627–632. doi: 10.1016/0022-1910(72)90190-4. - DOI
-
- Dartnall HJA. Photosensitivity. In: Dartnall HJA, editor. Handbook of sensory physiology, vol VII/1. Berlin: Springer; 1972. pp. 122–145.
-
- Dempster J (2001) The laboratory computer: a guide for neuroscientists and physiologists, Academic Press, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
