

The Sec14 superfamily and mechanisms for crosstalk between lipid metabolism and lipid signaling
- PMID: 19926291
- PMCID: PMC2834860
- DOI: 10.1016/j.tibs.2009.10.008
The Sec14 superfamily and mechanisms for crosstalk between lipid metabolism and lipid signaling
Abstract
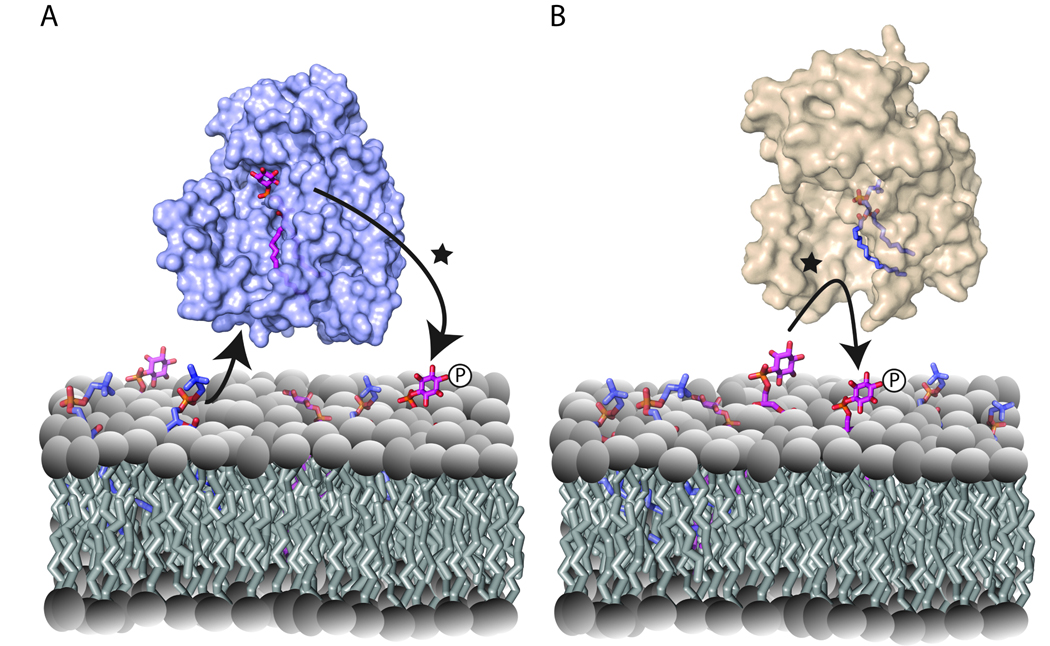
Lipid signaling pathways define central mechanisms for cellular regulation. Productive lipid signaling requires an orchestrated coupling between lipid metabolism, lipid organization and the action of protein machines that execute appropriate downstream reactions. Using membrane trafficking control as primary context, we explore the idea that the Sec14-protein superfamily defines a set of modules engineered for the sensing of specific aspects of lipid metabolism and subsequent transduction of 'sensing' information to a phosphoinositide-driven 'execution phase'. In this manner, the Sec14 superfamily connects diverse territories of the lipid metabolome with phosphoinositide signaling in a productive 'crosstalk' between these two systems. Mechanisms of crosstalk, by which non-enzymatic proteins integrate metabolic cues with the action of interfacial enzymes, represent unappreciated regulatory themes in lipid signaling.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures
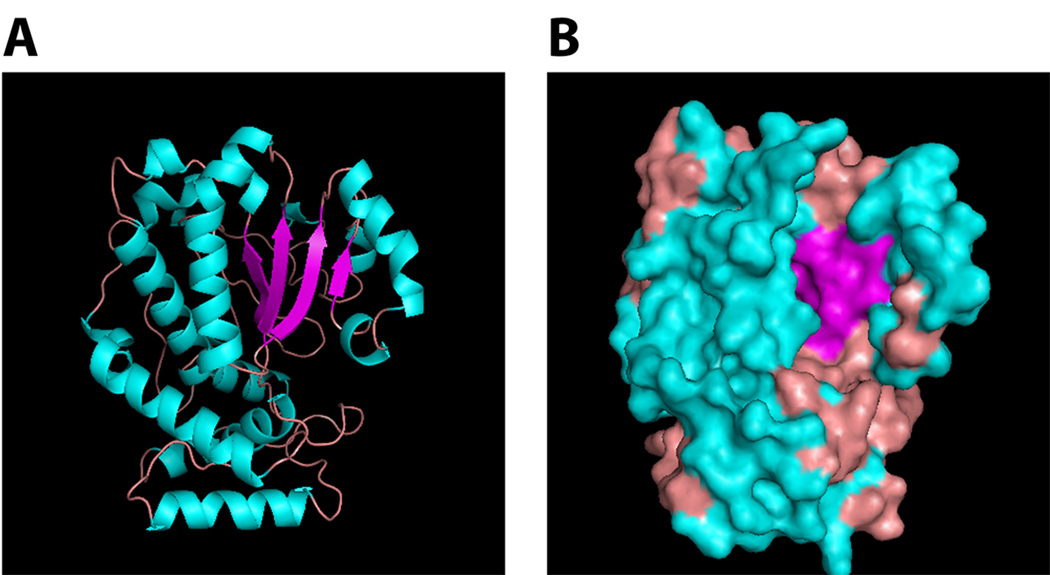
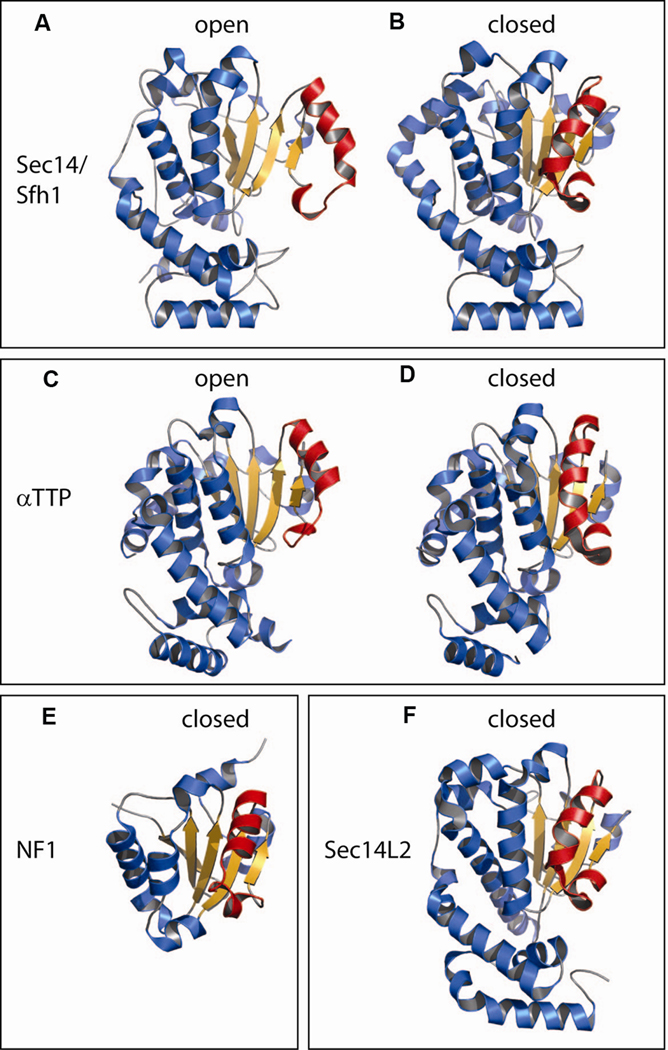
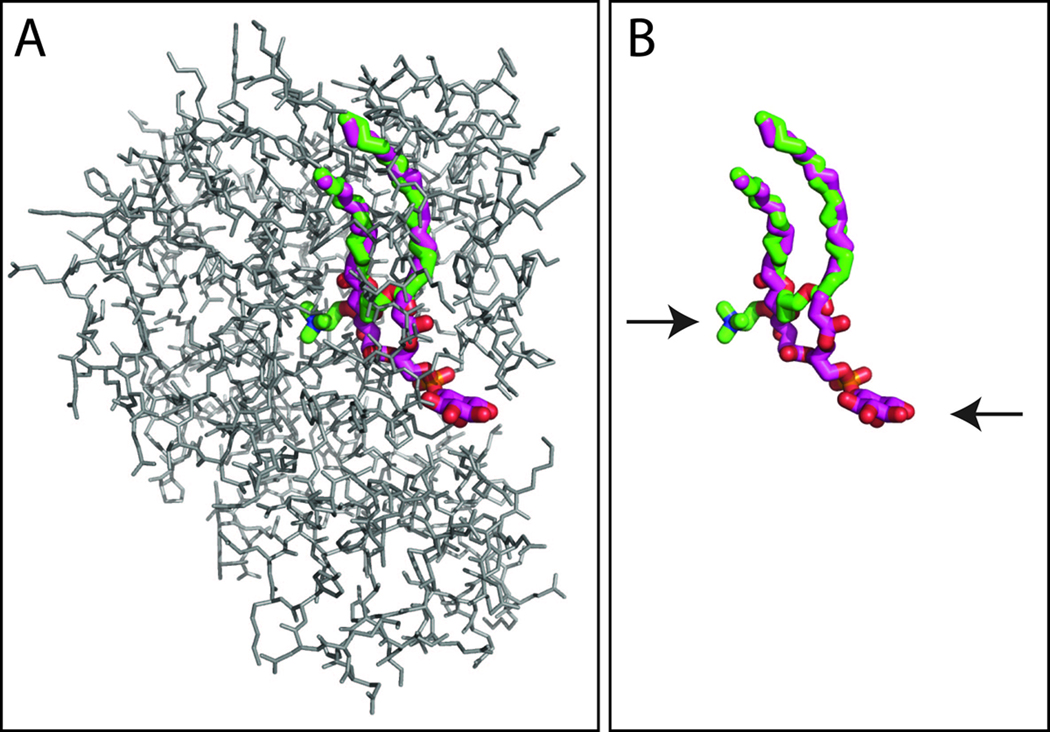
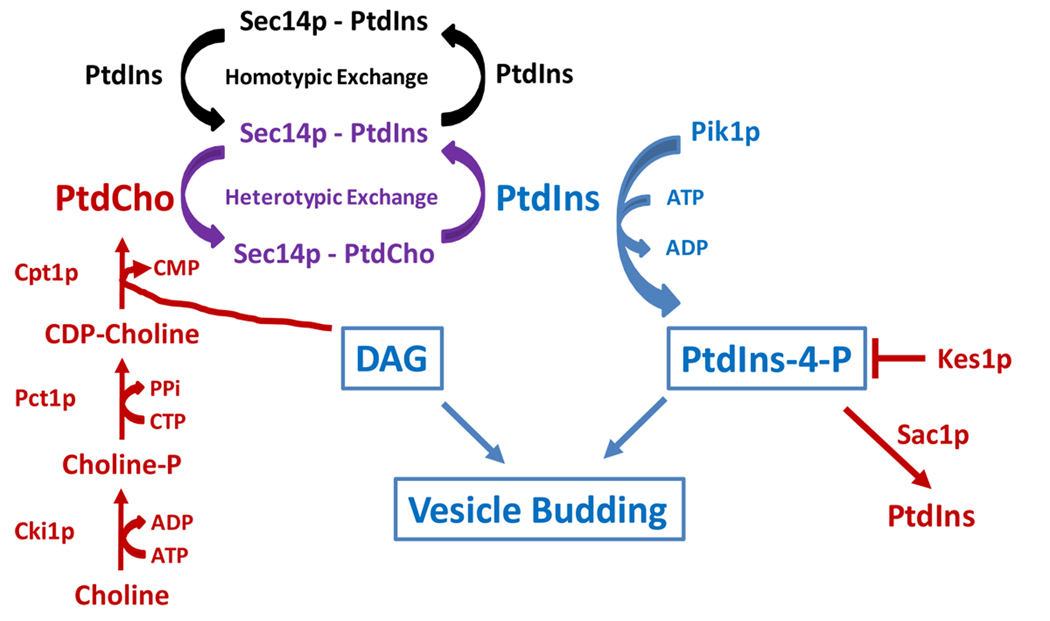
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
