

A progeria mutation reveals functions for lamin A in nuclear assembly, architecture, and chromosome organization
- PMID: 19926845
- PMCID: PMC2779830
- DOI: 10.1073/pnas.0911895106
A progeria mutation reveals functions for lamin A in nuclear assembly, architecture, and chromosome organization
Abstract
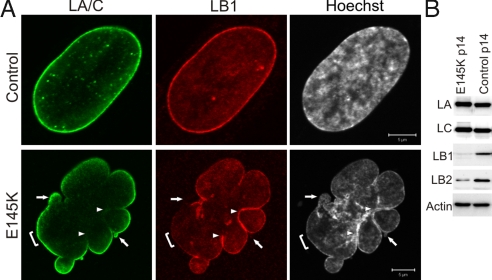
Numerous mutations in the human A-type lamin gene (LMNA) cause the premature aging disease, progeria. Some of these are located in the alpha-helical central rod domain required for the polymerization of the nuclear lamins into higher order structures. Patient cells with a mutation in this domain, 433G>A (E145K) show severely lobulated nuclei, a separation of the A- and B-type lamins, alterations in pericentric heterochromatin, abnormally clustered centromeres, and mislocalized telomeres. The induction of lobulations and the clustering of centromeres originate during postmitotic nuclear assembly in daughter cells and this early G1 configuration of chromosomes is retained throughout interphase. In vitro analyses of E145K-lamin A show severe defects in the assembly of protofilaments into higher order lamin structures. The results show that this central rod domain mutation affects nuclear architecture in a fashion distinctly different from the changes found in the most common form of progeria caused by the expression of LADelta50/progerin. The study also emphasizes the importance of lamins in nuclear assembly and chromatin organization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
