

Live imaging of rapid chromosome movements in meiotic prophase I in maize
- PMID: 19926853
- PMCID: PMC2791616
- DOI: 10.1073/pnas.0906498106
Live imaging of rapid chromosome movements in meiotic prophase I in maize
Abstract
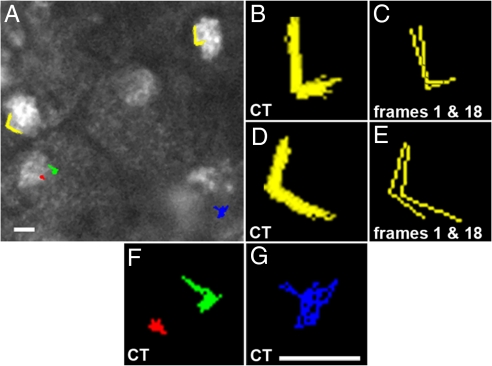
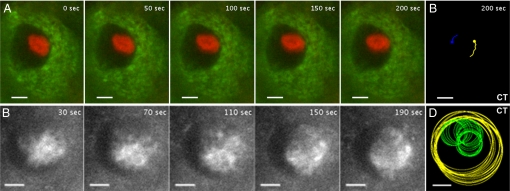
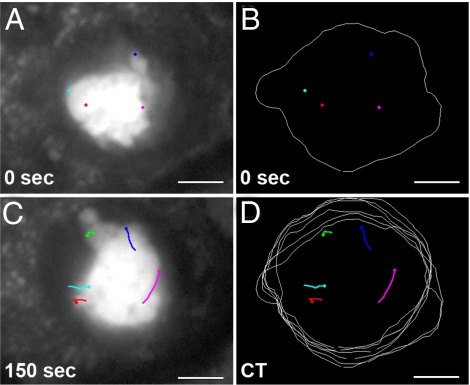
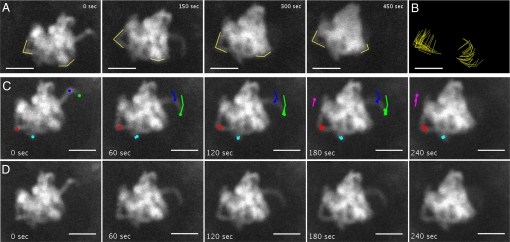
The ability of chromosomes to move across the nuclear space is essential for the reorganization of the nucleus that takes place in early meiotic prophase. Chromosome dynamics of prophase I have been studied in budding and fission yeasts, but little is known about this process in higher eukaryotes, where genomes and chromosomes are much larger and meiosis takes a longer time to complete. This knowledge gap has been mainly caused by difficulties in culturing isolated live meiocytes of multicellular eukaryotes. To study the nuclear dynamics during meiotic prophase in maize, we established a system to observe live meiocytes inside intact anthers. We found that maize chromosomes exhibited extremely dynamic and complex motility in zygonema and pachynema. The movement patterns differed dramatically between the two stages. Chromosome movements included rotations of the entire chromatin and movements of individual chromosome segments, which were mostly telomere-led. Chromosome motility was coincident with dynamic deformations of the nuclear envelope. Both, chromosome and nuclear envelope motility depended on actin microfilaments as well as tubulin. The complexity of the nuclear movements implies that several different mechanisms affect chromosome motility in early meiotic prophase in maize. We propose that the vigorous nuclear motility provides a mechanism for homologous loci to find each other during zygonema.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Zickler D, Kleckner N. The leptotene-zygotene transition of meiosis. Annu Rev Genet. 1998;32:619–697. - PubMed
-
- Dawe RK, Sedat JW, Agard DA, Cande WZ. Meiotic chromosome pairing in maize is associated with a novel chromatin organization. Cell. 1994;76:901–912. - PubMed
-
- Harper L, Golubovskaya I, Cande WZ. A bouquet of chromosomes. J Cell Sci. 2004;117:4025–4032. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
