

Combinations of genetic mutations in the adult neural stem cell compartment determine brain tumour phenotypes
- PMID: 19927122
- PMCID: PMC2808375
- DOI: 10.1038/emboj.2009.327
Combinations of genetic mutations in the adult neural stem cell compartment determine brain tumour phenotypes
Abstract
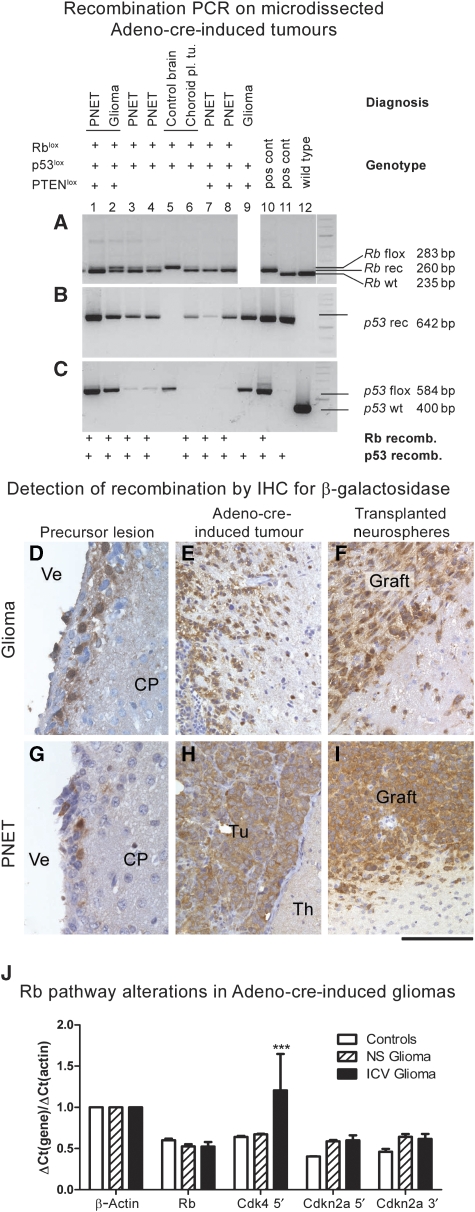
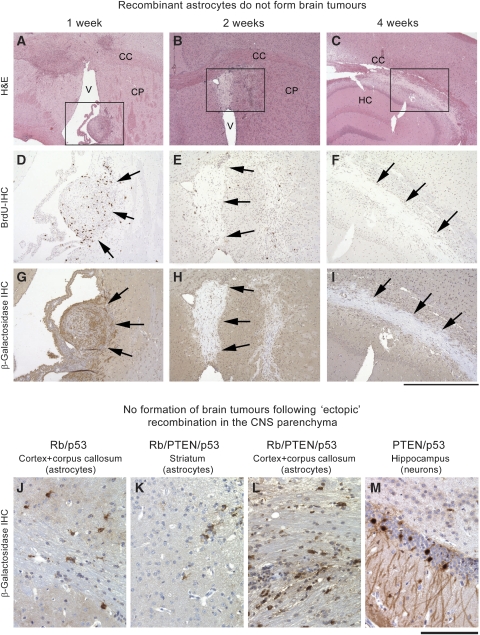
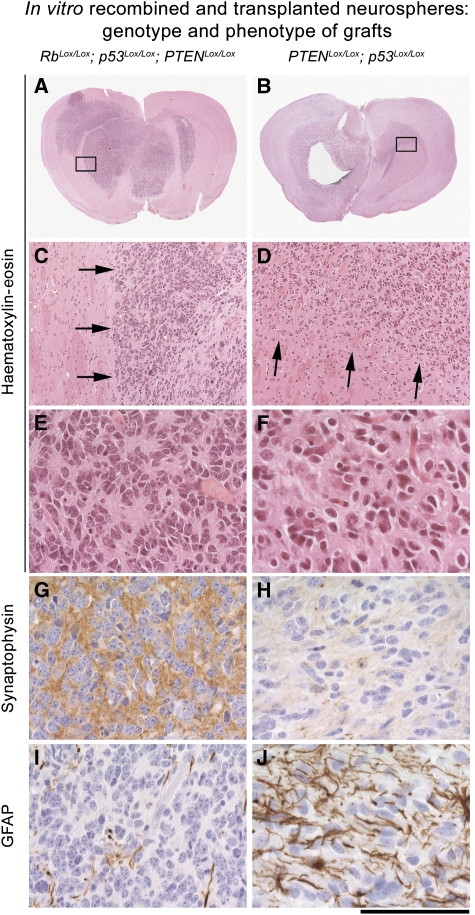
It has been suggested that intrinsic brain tumours originate from a neural stem/progenitor cell population in the subventricular zone of the post-natal brain. However, the influence of the initial genetic mutation on the phenotype as well as the contribution of mature astrocytes to the formation of brain tumours is still not understood. We deleted Rb/p53, Rb/p53/PTEN or PTEN/p53 in adult subventricular stem cells; in ectopically neurografted stem cells; in mature parenchymal astrocytes and in transplanted astrocytes. We found that only stem cells, but not astrocytes, gave rise to brain tumours, independent of their location. This suggests a cell autonomous mechanism that enables stem cells to generate brain tumours, whereas mature astrocytes do not form brain tumours in adults. Recombination of PTEN/p53 gave rise to gliomas whereas deletion of Rb/p53 or Rb/p53/PTEN generated primitive neuroectodermal tumours (PNET), indicating an important role of an initial Rb loss in driving the PNET phenotype. Our study underlines an important role of stem cells and the relevance of initial genetic mutations in the pathogenesis and phenotype of brain tumours.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
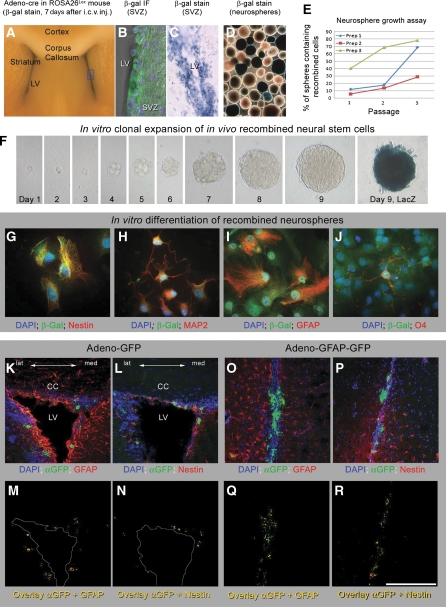
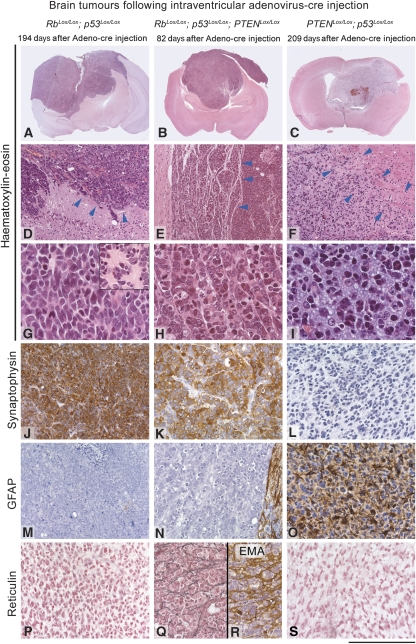
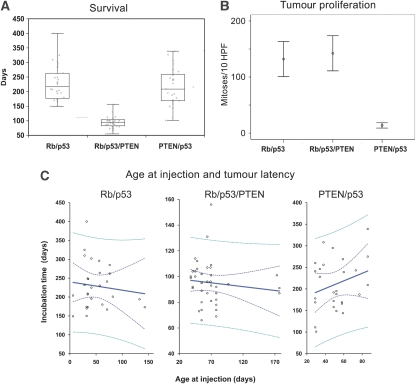
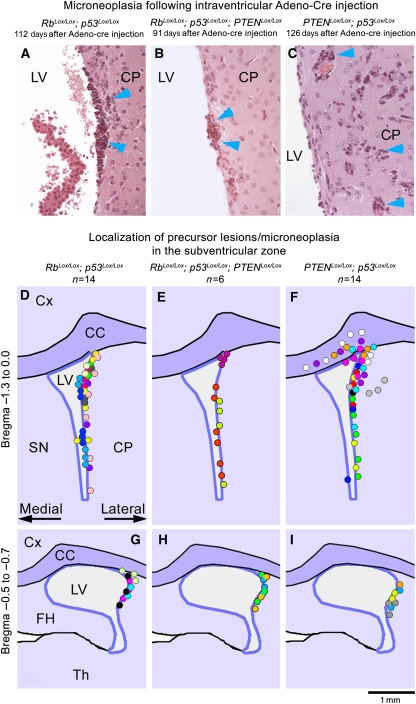
References
-
- Alvarez-Buylla A, Kohwi M, Nguyen TM, Merkle FT (2008) The heterogeneity of adult neural stem cells and the emerging complexity of their niche. Cold Spring Harb Symp Quant Biol 73: 357–365 - PubMed
-
- Alvarez-Buylla A, Temple S (1998) Stem cells in the developing and adult nervous system. J Neurobiol 36: 105–110 - PubMed
-
- Bachoo RM, Maher EA, Ligon KL, Sharpless NE, Chan SS, You MJ, Tang Y, DeFrances J, Stover E, Weissleder R, Rowitch DH, Louis DN, DePinho RA (2002) Epidermal growth factor receptor and Ink4a/Arf: convergent mechanisms governing terminal differentiation and transformation along the neural stem cell to astrocyte axis. Cancer Cell 1: 269–277 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
