

Ca(2+) channels on the move
- PMID: 19928968
- PMCID: PMC2797372
- DOI: 10.1021/bi901739t
Ca(2+) channels on the move
Erratum in
- Biochemistry. 2010 Jun 22;49(24):5082
Abstract
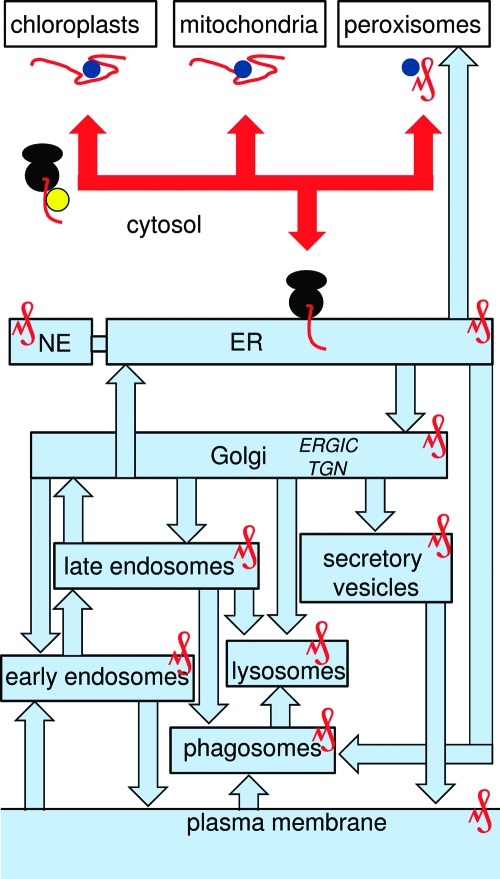
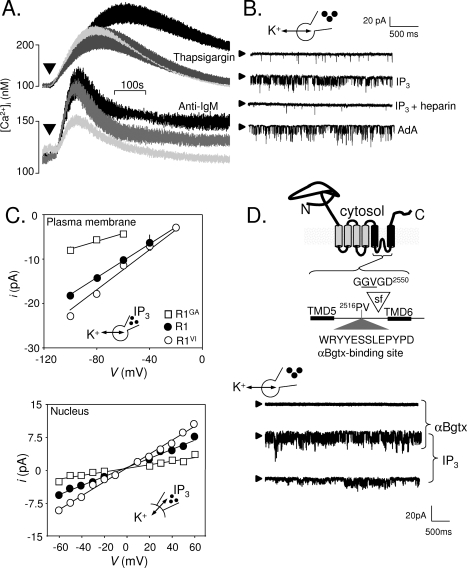
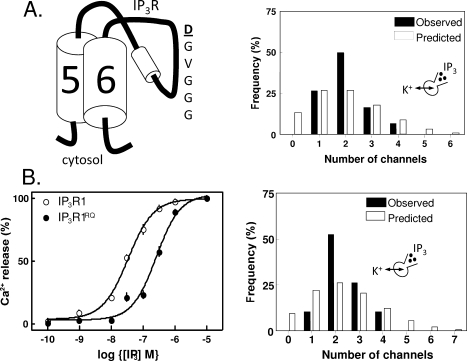
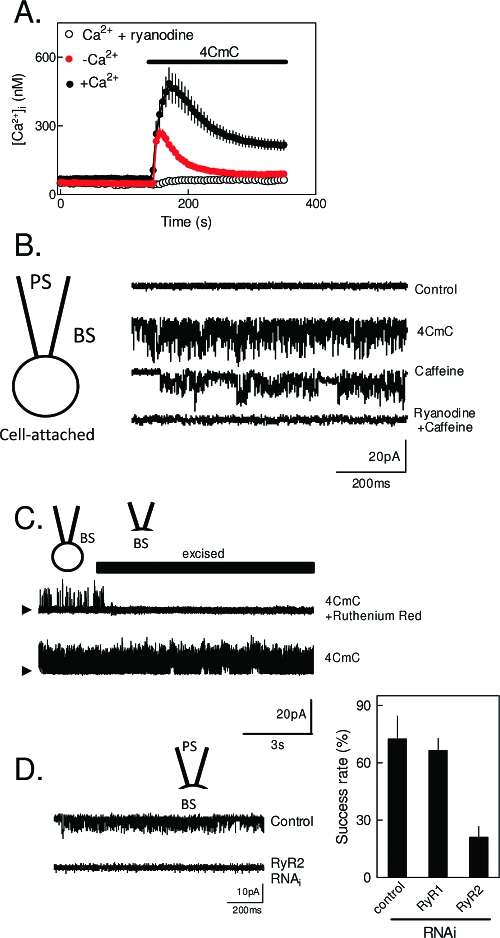
The versatility of Ca(2+) as an intracellular messenger derives largely from the spatial organization of cytosolic Ca(2+) signals, most of which are generated by regulated openings of Ca(2+)-permeable channels. Most Ca(2+) channels are expressed in the plasma membrane (PM). Others, including the almost ubiquitous inositol 1,4,5-trisphosphate receptors (IP(3)R) and their relatives, the ryanodine receptors (RyR), are predominantly expressed in membranes of the sarcoplasmic or endoplasmic reticulum (ER). Targeting of these channels to appropriate destinations underpins their ability to generate spatially organized Ca(2+) signals. All Ca(2+) channels begin life in the cytosol, and the vast majority are then functionally assembled in the ER, where they may either remain or be dispatched to other membranes. Here, by means of selective examples, we review two issues related to this trafficking of Ca(2+) channels via the ER. How do cells avoid wayward activity of Ca(2+) channels in transit as they pass from the ER via other membranes to their final destination? How and why do some cells express small numbers of the archetypal intracellular Ca(2+) channels, IP(3)R and RyR, in the PM?
Figures
References
-
- Berridge M. J.; Bootman M. D.; Roderick H. L. (2003) Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 4, 517–529. - PubMed
-
- Berridge M. J.; Lipp P.; Bootman M. D. (2000) The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 1, 11–21. - PubMed
-
- Xiong T. C.; Bourque S.; Lecourieux D.; Amelot N.; Grat S.; Briere C.; Mazars C.; Pugin A.; Ranjeva R. (2006) Calcium signaling in plant cell organelles delimited by a double membrane. Biochim. Biophys. Acta 1763, 1209–1215. - PubMed
-
- Kirichok Y.; Krapavinsky G.; Clapham D. E. (2004) The mitochondrial calcium uniporter is a highly selective ion channel. Nature 427, 360–364. - PubMed
-
- Toyoshima C. (2008) Structural aspects of ion pumping by Ca2+-ATPase of sarcoplasmic reticulum. Arch. Biochem. Biophys. 476, 3–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
