

Relationships between cycling hypoxia, HIF-1, angiogenesis and oxidative stress
- PMID: 19929412
- PMCID: PMC2790140
- DOI: 10.1667/RR1926.1
Relationships between cycling hypoxia, HIF-1, angiogenesis and oxidative stress
Abstract
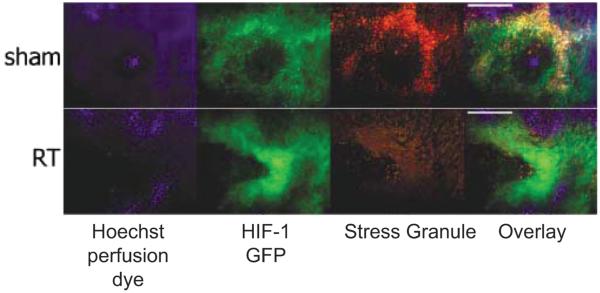
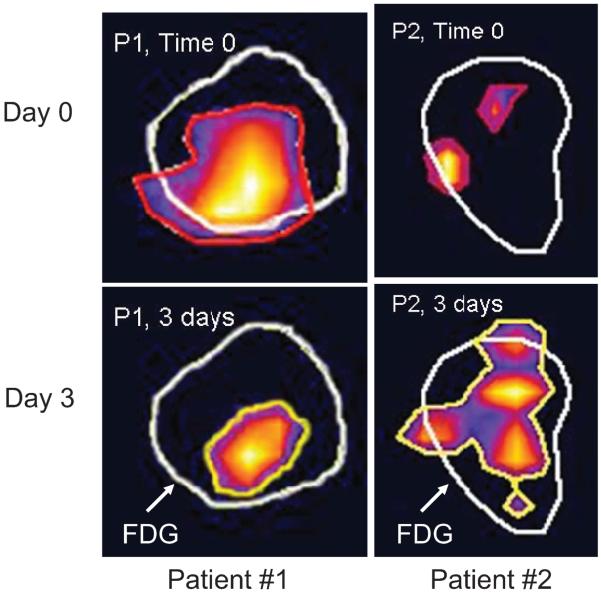
This Failla Lecture focused on the inter-relationships between tumor angiogenesis, HIF-1 expression and radiotherapy responses. A common thread that bonds all of these factors together is microenvironmental stress caused by reactive oxygen and nitrogen species formed during tumor growth and angiogenesis or in response to cytotoxic treatment. In this review we focus on one aspect of the crossroad between oxidative stress and angiogenesis, namely cycling hypoxia. Understanding of the relative importance of this feature of the tumor microenvironment has recently expanded; it influences tumor biology in ways that are separate from chronic hypoxia. Cycling hypoxia can influence angiogenesis, treatment responses and metastatic behavior. It represents an important and relatively less well understood feature of tumor biology that requires additional research.
Figures
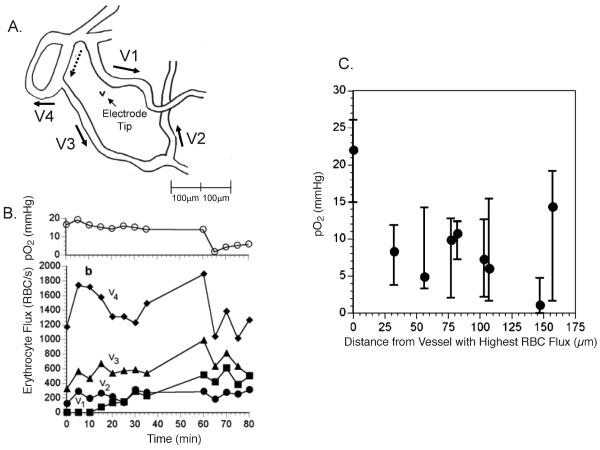
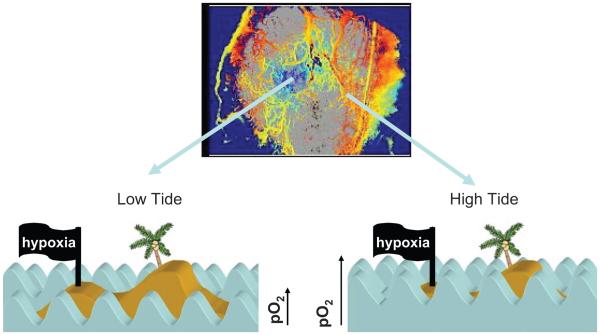
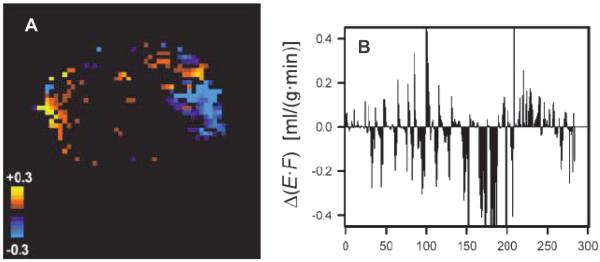
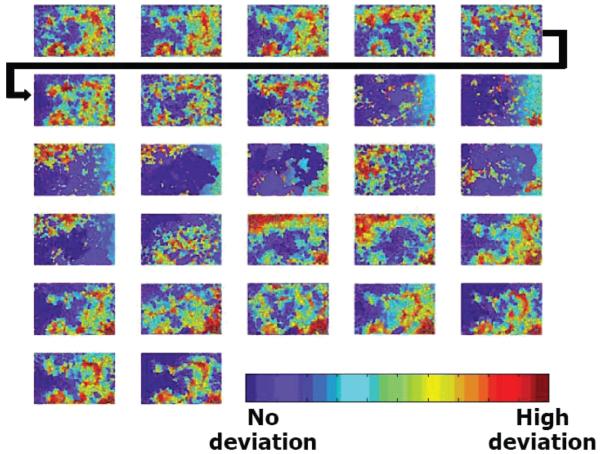
References
-
- Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671–674. - PubMed
-
- Ferrara N, Davis-Smyth T. The biology of vascular endothelial growth factor. Endocr. Rev. 1997;18:4–25. - PubMed
-
- Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J. 1999;13:9–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
