

Dissection of platelet and myeloid cell defects by conditional targeting of the beta3-integrin subunit
- PMID: 19933310
- PMCID: PMC2845430
- DOI: 10.1096/fj.09-138420
Dissection of platelet and myeloid cell defects by conditional targeting of the beta3-integrin subunit
Abstract
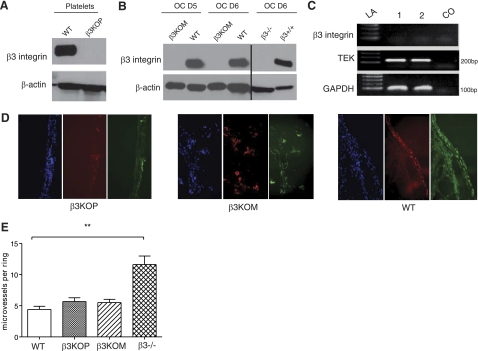
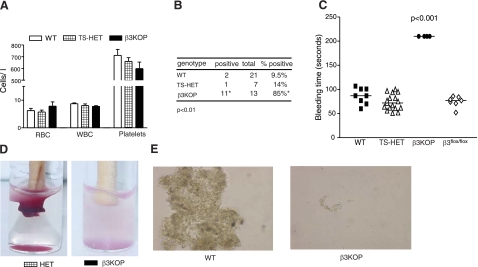
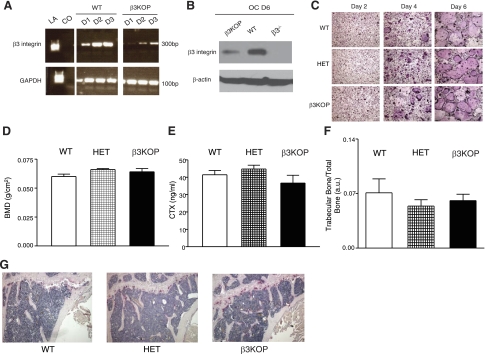
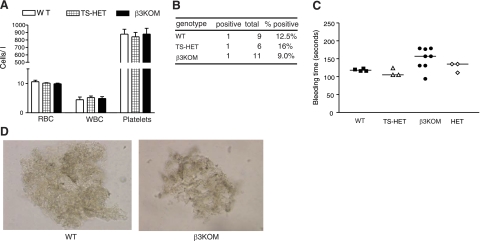
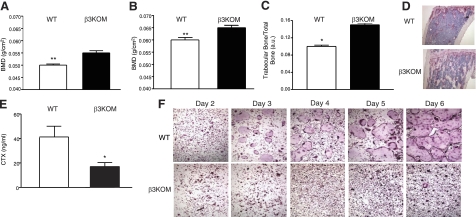
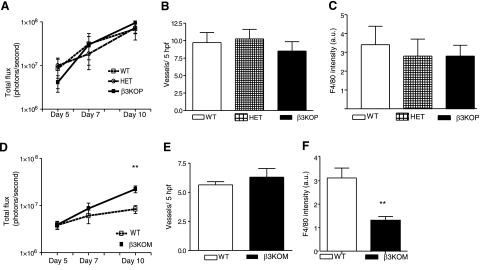
The purpose of this work was to determine platelet and myeloid cell-specific requirements for beta3-containing integrins in hemostasis, bone resorption, and tumor growth. LoxP-flanked mice were generated to study the conditional deletion of beta3-integrin in platelets [knockout in platelets (KOP)] and myeloid cells [knockout in myeloid (KOM)]. Using the beta3KOP and beta3KOM strains of mice, we studied the role of beta3-integrin in hemostasis, bone resorption, and subcutaneous tumor growth. Tissue-specific deletion of platelet beta3-integrins in beta3KOP mice did not affect bone mass but resulted in a severe bleeding phenotype. No growth difference of tumor xenografts or in neoangiogenesis were found in beta3KOP mice, in contrast to the defects observed in germline beta3(-/-) mice. Conditional deletion of myeloid beta3-integrins in beta3KOM mice resulted in osteopetrosis but had no effect on hemostasis or mortality. Tumor growth in beta3KOM mice was increased and accompanied by decreased macrophage infiltration, without increase in blood vessel number. Platelet beta3-integrin deficiency was sufficient to disrupt hemostasis but had no effect on bone mass or tumor growth. Myeloid-specific beta3-integrin deletion was sufficient to perturb bone mass and enhance tumor growth due to reduced macrophage infiltration in the tumors. These results suggest that beta3-integrins have cell-specific roles in complex biological processes.-Morgan, E. A., Schneider, J. G., Baroni, T. E., Uluçkan, O., Heller, E., Hurchla, M. A., Deng, H., Floyd, D., Berdy, A., Prior, J. L., Piwnica-Worms, D., Teitelbaum, S. L., Ross, F. P., Weilbaecher, K. N. Dissection of platelet and myeloid cell defects by conditional targeting of the beta3-integrin subunit.
Figures
Similar articles
-
Critical role of beta3 integrin in experimental postmenopausal osteoporosis.J Bone Miner Res. 2005 Dec;20(12):2116-23. doi: 10.1359/JBMR.050724. Epub 2005 Jul 25. J Bone Miner Res. 2005. PMID: 16294265
-
Cooperation Between Platelet β1 and β3 Integrins in the Arrest of Bleeding Under Inflammatory Conditions in Mice-Brief Report.Arterioscler Thromb Vasc Biol. 2024 Oct;44(10):2213-2222. doi: 10.1161/ATVBAHA.124.321104. Epub 2024 Aug 15. Arterioscler Thromb Vasc Biol. 2024. PMID: 39145395
-
Talin is required for integrin-mediated platelet function in hemostasis and thrombosis.J Exp Med. 2007 Dec 24;204(13):3103-11. doi: 10.1084/jem.20071800. Epub 2007 Dec 17. J Exp Med. 2007. PMID: 18086863 Free PMC article.
-
Effects of thrombin on interactions between beta3-integrins and extracellular matrix in platelets and vascular cells.Arterioscler Thromb Vasc Biol. 2003 Nov 1;23(11):1971-8. doi: 10.1161/01.ATV.0000093470.51580.0F. Epub 2003 Aug 28. Arterioscler Thromb Vasc Biol. 2003. PMID: 12947018 Review.
-
The Possible Importance of β3 Integrins for Leukemogenesis and Chemoresistance in Acute Myeloid Leukemia.Int J Mol Sci. 2018 Jan 15;19(1):251. doi: 10.3390/ijms19010251. Int J Mol Sci. 2018. PMID: 29342970 Free PMC article. Review.
Cited by
-
Slow integrin-dependent migration organizes networks of tissue-resident mast cells.Nat Immunol. 2023 Jun;24(6):915-924. doi: 10.1038/s41590-023-01493-2. Epub 2023 Apr 20. Nat Immunol. 2023. PMID: 37081147 Free PMC article.
-
Integrin β3 in forebrain Emx1-expressing cells regulates repetitive self-grooming and sociability in mice.BMC Neurosci. 2022 Mar 5;23(1):12. doi: 10.1186/s12868-022-00691-2. BMC Neurosci. 2022. PMID: 35247972 Free PMC article.
-
Integrin αvβ3 Limits Cytokine Production by Plasmacytoid Dendritic Cells and Restricts TLR-Driven Autoimmunity.J Immunol. 2024 Jun 1;212(11):1680-1692. doi: 10.4049/jimmunol.2300290. J Immunol. 2024. PMID: 38607278 Free PMC article.
-
Thrombospondin-1 regulates bone homeostasis through effects on bone matrix integrity and nitric oxide signaling in osteoclasts.J Bone Miner Res. 2015 Jan;30(1):106-15. doi: 10.1002/jbmr.2308. J Bone Miner Res. 2015. PMID: 25042340 Free PMC article.
-
The age of bone marrow dictates the clonality of smooth muscle-derived cells in atherosclerotic plaques.Nat Aging. 2023 Jan;3(1):64-81. doi: 10.1038/s43587-022-00342-5. Epub 2023 Jan 9. Nat Aging. 2023. PMID: 36743663 Free PMC article.
References
-
- Phillips D R, Charo I F, Scarborough R M. GPIIb-IIIa: the responsive integrin. Cell. 1991;65:359–362. - PubMed
-
- Hynes R O. Integrins: versatility, modulation, and signaling in cell adhesion. Cell. 1992;69:11–25. - PubMed
-
- Schwartz M A, Schaller M D, Ginsberg M H. Integrins: emerging paradigms of signal transduction. Annu Rev Cell Dev Biol. 1995;11:549–599. - PubMed
-
- Hynes R O. Integrins: a family of cell surface receptors. Cell. 1987;48:549–554. - PubMed
-
- Brooks P C, Clark R A, Cheresh D A. Requirement of vascular integrin alpha v beta 3 for angiogenesis. Science. 1994;264:569–571. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AR032788/AR/NIAMS NIH HHS/United States
- R01 AR046523/AR/NIAMS NIH HHS/United States
- R37 AR046523/AR/NIAMS NIH HHS/United States
- AR046852/AR/NIAMS NIH HHS/United States
- AR046523/AR/NIAMS NIH HHS/United States
- T32 GM007200/GM/NIGMS NIH HHS/United States
- T32 GM7200/GM/NIGMS NIH HHS/United States
- R01 AR032788/AR/NIAMS NIH HHS/United States
- P50 CA94056/CA/NCI NIH HHS/United States
- P30 AR057235/AR/NIAMS NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- DK56341/DK/NIDDK NIH HHS/United States
- NIHR0152152/PHS HHS/United States
- P50 CA094056/CA/NCI NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- R01 AR046852/AR/NIAMS NIH HHS/United States
- R56 CA097250/CA/NCI NIH HHS/United States
- R01 CA097250/CA/NCI NIH HHS/United States
- P30 DK52574/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
