

The ArcBA two-component system of Escherichia coli is regulated by the redox state of both the ubiquinone and the menaquinone pool
- PMID: 19933363
- PMCID: PMC2812447
- DOI: 10.1128/JB.01156-09
The ArcBA two-component system of Escherichia coli is regulated by the redox state of both the ubiquinone and the menaquinone pool
Abstract
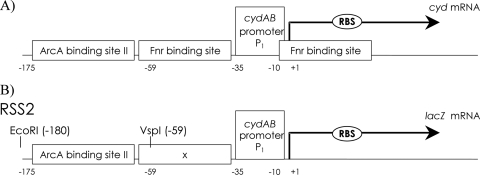
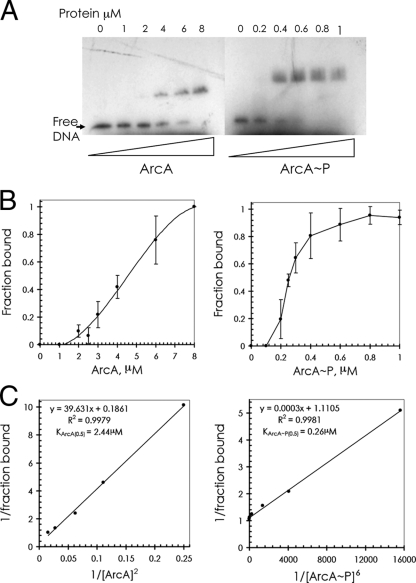
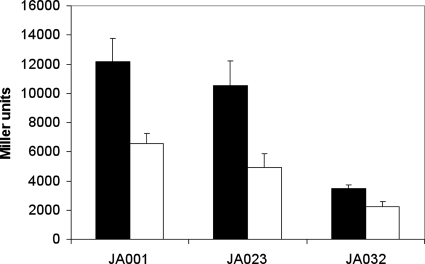
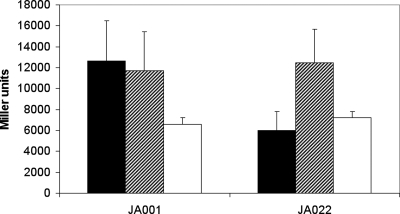
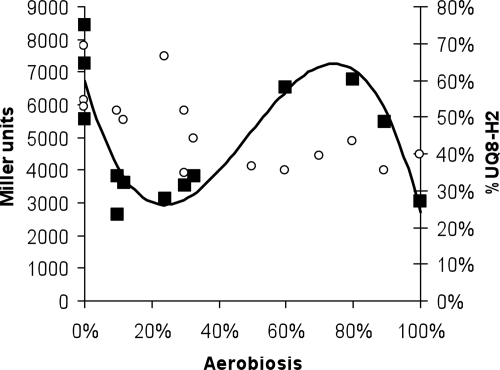
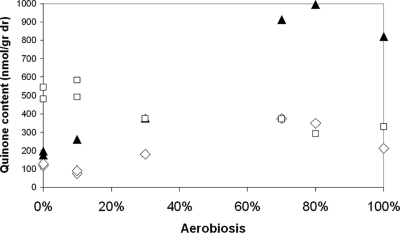
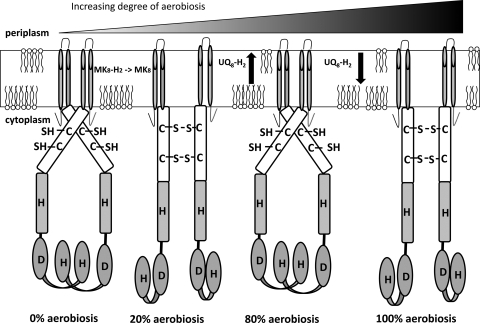
ArcBA is a two-component regulatory system of Escherichia coli involved in sensing oxygen availability and the concomitant transcriptional regulation of oxidative and fermentative catabolism. Based on in vitro data, it has been postulated that the redox state of the ubiquinone pool is the determinant for ArcB kinase activity. Here we report on the in vivo regulation of ArcB activation, as determined using a lacZ reporter specifically responsive to phosphorylated ArcA. Our results indicate that upon deletion of a ubiquinone biosynthetic enzyme, regulation of ArcB in the anaerobic-aerobic transition is not affected. In contrast, interference with menaquinone biosynthesis leads to inactivation of ArcB during anaerobic growth; this phenotype is fully rescued by addition of a menaquinone precursor. This clearly demonstrates that the menaquinones play a major role in ArcB activation. ArcB shows a complex pattern of regulation when E. coli is titrated through the entire aerobiosis range; ArcB is activated under anaerobic and subaerobic conditions and is much less active under fully aerobic and microaerobic conditions. Furthermore, there is no correlation between ArcB activation and the redox state of the ubiquinone pool, but there is a restricted correlation between the total cellular ubiquinone content and ArcB activity due to the considerable increase in the size of the ubiquinone pool with increasing degrees of aerobiosis. These results lead to the working hypothesis that the in vivo activity of ArcB in E. coli is modulated by the redox state of the menaquinone pool and that the ubiquinone/ubiquinol ratio in vivo surely is not the only determinant of ArcB activity.
Figures
References
-
- Alexeeva, S. 2000. Molecular physiology of responses to oxygen in Escherichia coli. Ph.D. thesis. University of Amsterdam, Amsterdam, The Netherlands.
-
- Bekker, M., G. Kramer, A. F. Hartog, M. J. Wagner, C. G. de Koster, K. J. Hellingwerf, and M. J. de Mattos. 2007. Changes in the redox state and composition of the quinone pool of Escherichia coli during aerobic batch-culture growth. Microbiology 153:1974-1980. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
