

Transitions in oral and intestinal microflora composition and innate immune receptor-dependent stimulation during mouse development
- PMID: 19933833
- PMCID: PMC2812188
- DOI: 10.1128/IAI.01043-09
Transitions in oral and intestinal microflora composition and innate immune receptor-dependent stimulation during mouse development
Abstract
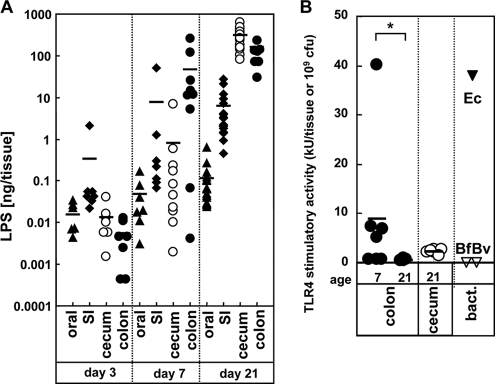
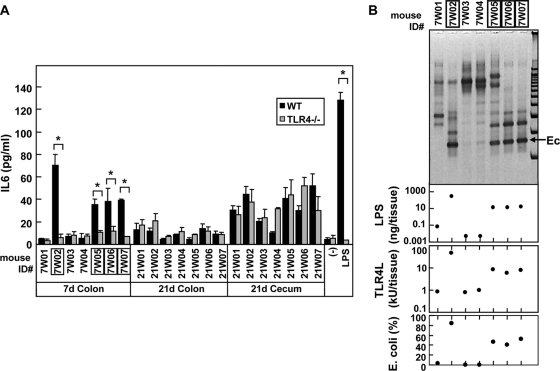
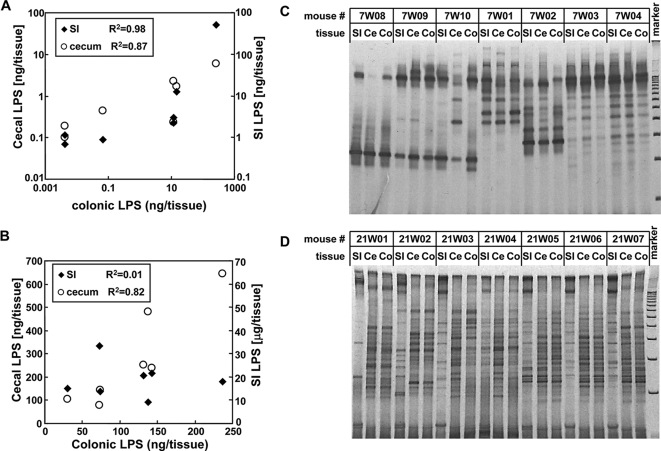
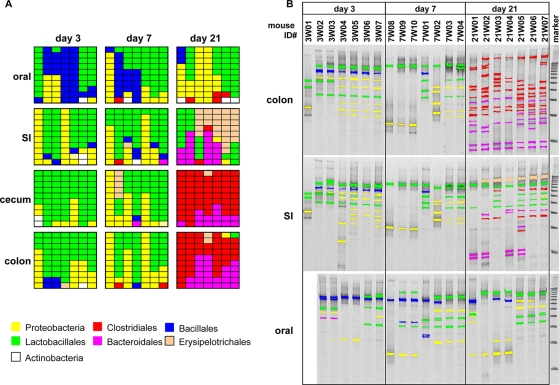
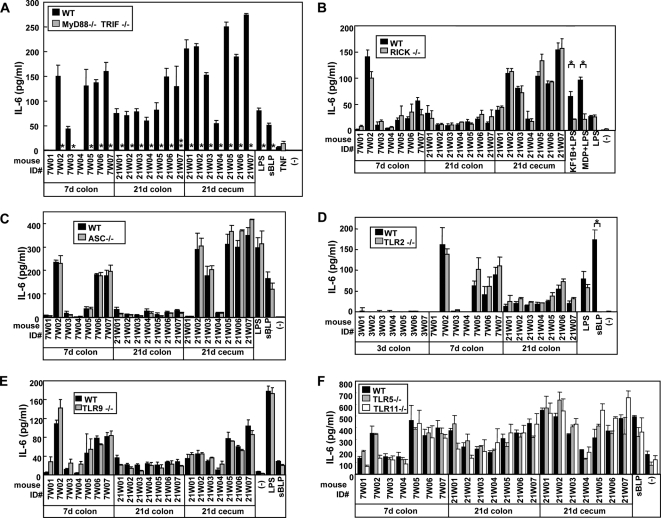
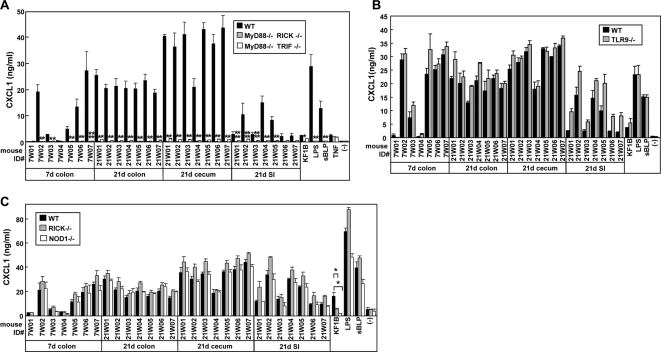
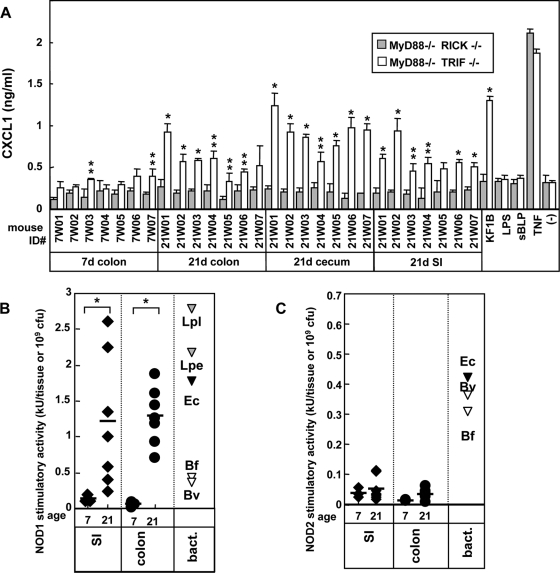
Commensal bacteria possess immunostimulatory activities that can modulate host responses to affect development and homeostasis in the intestine. However, how different populations of resident bacteria stimulate the immune system remains largely unknown. We characterized here the ability of intestinal and oral microflora to stimulate individual pattern recognition receptors (PRRs) in bone marrow-derived macrophages and mesothelial cells. The intestinal but not oral microflora elicited age- and cell type-specific immunostimulation. The immunostimulatory activity of the intestinal microflora varied among individual mice but was largely mediated via Toll-like receptor 4 (TLR4) during breast-feeding, whereas it became TLR4 independent after weaning. This transition was associated with a change from a microflora rich in TLR4-stimulatory proteobacteria to one dominated by Bacteroidales and/or Clostridiales that poorly stimulate TLR4. The major stimulatory activity of the intestinal microflora was still intact in NOD1-, NOD2-, TLR2-, TLR4-, TLR5-, TLR9-, TLR11-, ASC-, or RICK-deficient cells but still relied on the adaptor MyD88. These studies demonstrate a transition in the intestinal microflora accompanied by a dynamic change of its ability to stimulate different PRRs which control intestinal homeostasis.
Figures

References
-
- Reference deleted.
-
- Artis, D. 2008. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 8:411-420. - PubMed
-
- Atarashi, K., J. Nishimura, T. Shima, Y. Umesaki, M. Yamamoto, M. Onoue, H. Yagita, N. Ishii, R. Evans, K. Honda, and K. Takeda. 2008. ATP drives lamina propriaT(H)17 cell differentiation. Nature 455:808-812. - PubMed
-
- Bartosch, S., A. Fite, G. T. Macfarlane, and M. E. McMurdo. 2004. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl. Environ. Microbiol. 70:3575-3581. - PMC - PubMed
-
- Bouskra, D., C. Brézillon, M. Bérard, C. Werts, R. Varona, I. G. Boneca, and G. Eberl. 2008. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature 456:507-510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
