

In a murine tuberculosis model, the absence of homeostatic chemokines delays granuloma formation and protective immunity
- PMID: 19933855
- PMCID: PMC2799945
- DOI: 10.4049/jimmunol.0901937
In a murine tuberculosis model, the absence of homeostatic chemokines delays granuloma formation and protective immunity
Abstract
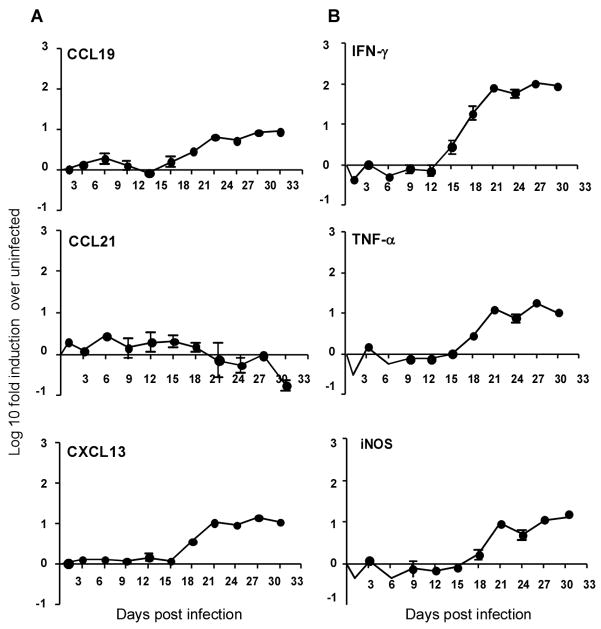
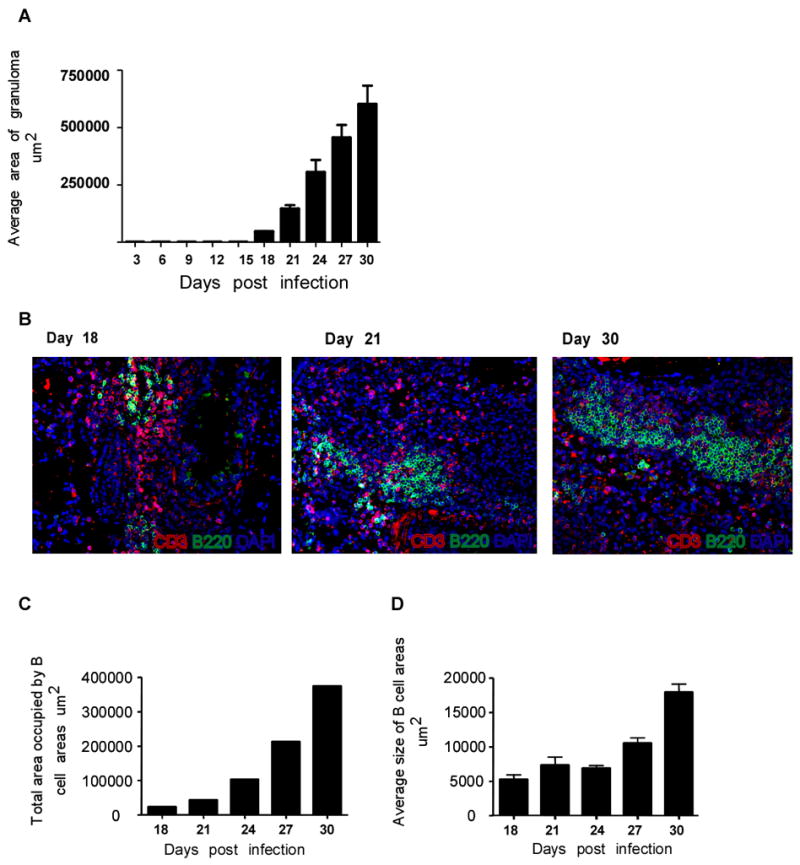
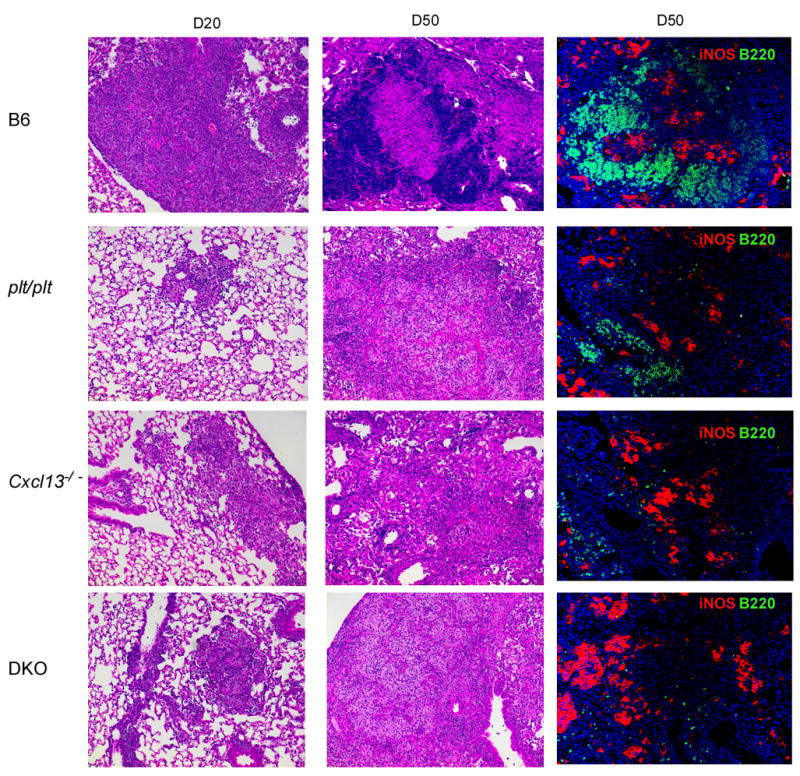
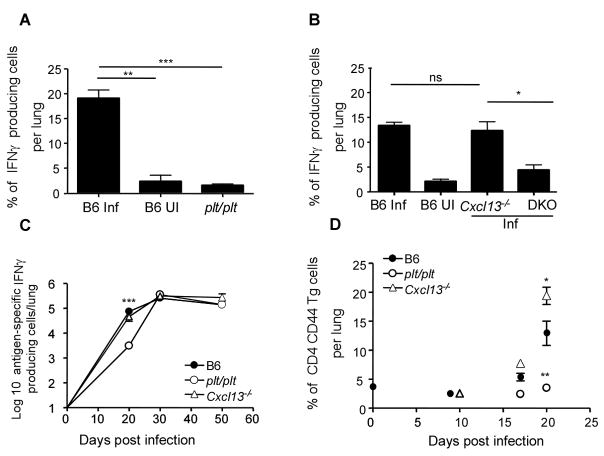
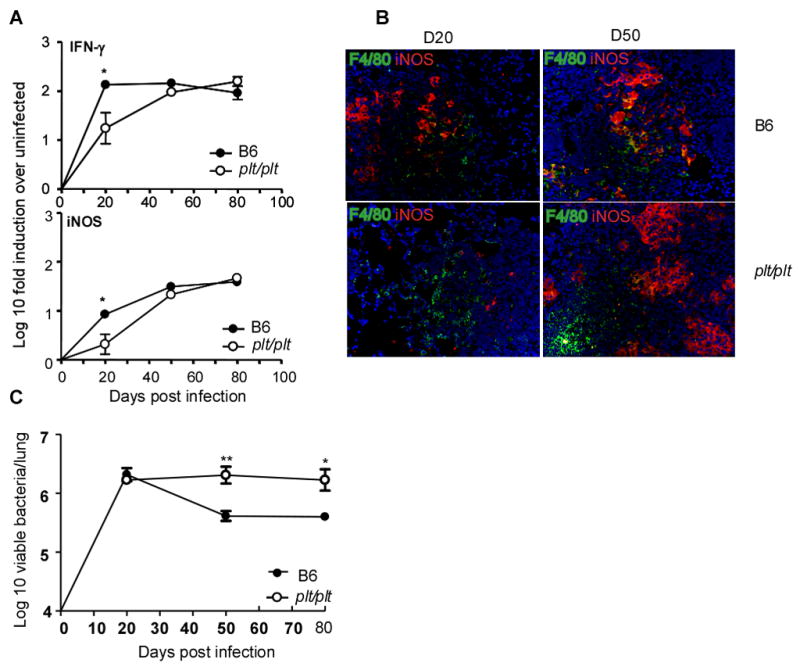
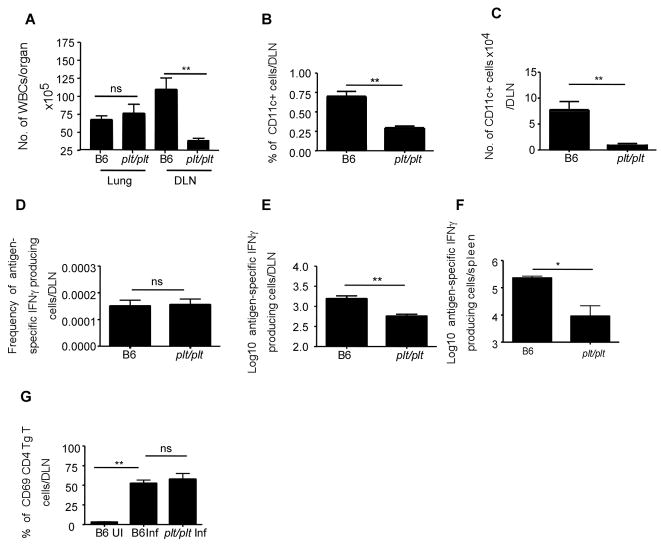
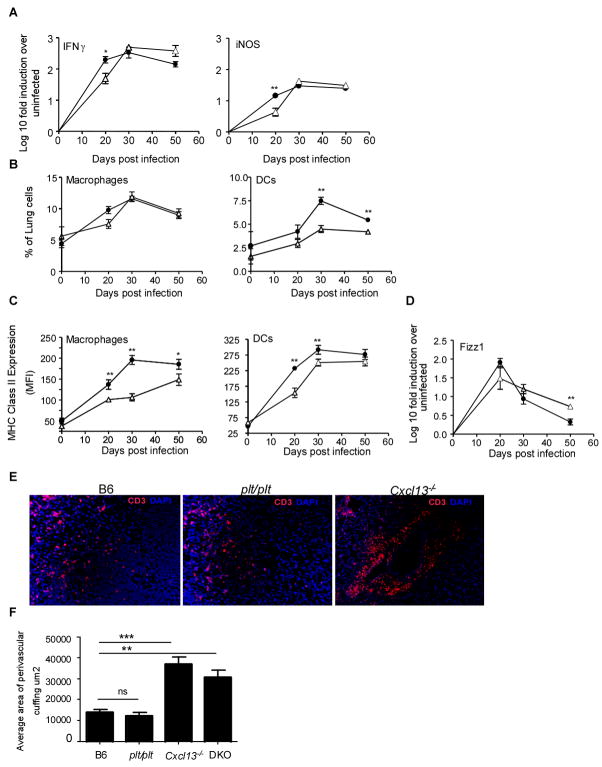
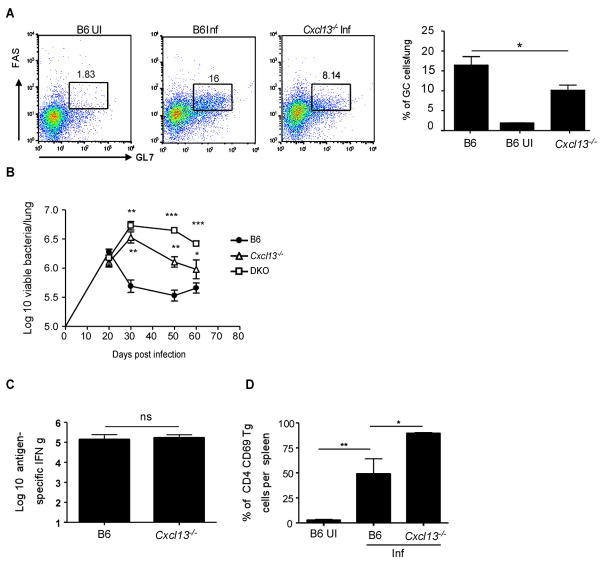
Mycobacterium tuberculosis infection (Mtb) results in the generation of protective cellular immunity and formation of granulomatous structures in the lung. CXCL13, CCL21, and CCL19 are constitutively expressed in the secondary lymphoid organs and play a dominant role in the homing of lymphocytes and dendritic cells. Although it is known that dendritic cell transport of Mtb from the lung to the draining lymph node is dependent on CCL19/CCL21, we show in this study that CCL19/CCL21 is also important for the accumulation of Ag-specific IFN-gamma-producing T cells in the lung, development of the granuloma, and control of mycobacteria. Importantly, we also show that CXCL13 is not required for generation of IFN-gamma responses, but is essential for the spatial arrangement of lymphocytes within granulomas, optimal activation of phagocytes, and subsequent control of mycobacterial growth. Furthermore, we show that these chemokines are also induced in the lung during the early immune responses following pulmonary Mtb infection. These results demonstrate that homeostatic chemokines perform distinct functions that cooperate to mediate effective expression of immunity against Mtb infection.
Figures
References
-
- Flynn J, Chan J. Immunology of tuberculosis. Annual Review of Immunology. 2001;19:93–129. - PubMed
-
- Bean AG, Roach DR, Briscoe H, France MP, Korner H, Sedgwick JD, Britton WJ. Structural deficiencies in granuloma formation in TNF gene-targeted mice underlie the heightened susceptibility to aerosol Mycobacterium tuberculosis infection, which is not compensated for by lymphotoxin. J Immunol. 1999;162:3504–3511. - PubMed
-
- Cyster JG. Chemokines and cell migration in secondary lymphoid organs. Science. 1999;286:2098–2102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI-46530/AI/NIAID NIH HHS/United States
- P01 AI046530/AI/NIAID NIH HHS/United States
- 1K99/R00AI075106-01/AI/NIAID NIH HHS/United States
- AI-67723/AI/NIAID NIH HHS/United States
- R01 AI072689/AI/NIAID NIH HHS/United States
- AI073564/AI/NIAID NIH HHS/United States
- U54 AI057158/AI/NIAID NIH HHS/United States
- AI057158/AI/NIAID NIH HHS/United States
- AI072689/AI/NIAID NIH HHS/United States
- R00 AI075106/AI/NIAID NIH HHS/United States
- HL69409/HL/NHLBI NIH HHS/United States
- R01 HL069409/HL/NHLBI NIH HHS/United States
- R01 AI073564/AI/NIAID NIH HHS/United States
- K99 AI075106/AI/NIAID NIH HHS/United States
- R01 AI067723/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
