

Tumor progression locus 2 (Map3k8) is critical for host defense against Listeria monocytogenes and IL-1 beta production
- PMID: 19933865
- PMCID: PMC2946069
- DOI: 10.4049/jimmunol.0901336
Tumor progression locus 2 (Map3k8) is critical for host defense against Listeria monocytogenes and IL-1 beta production
Abstract
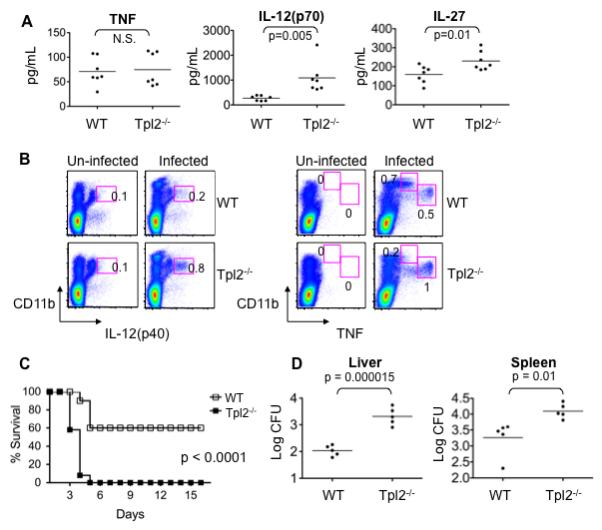
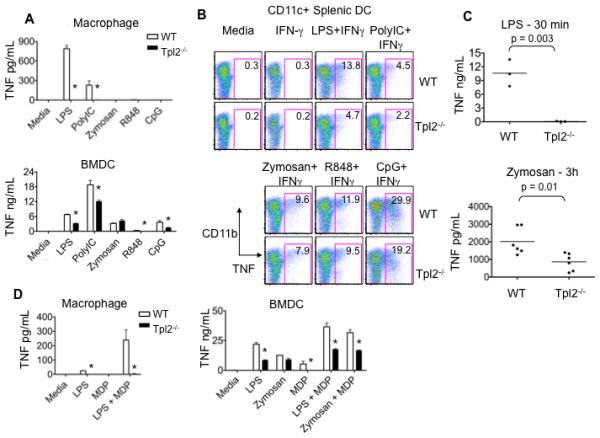
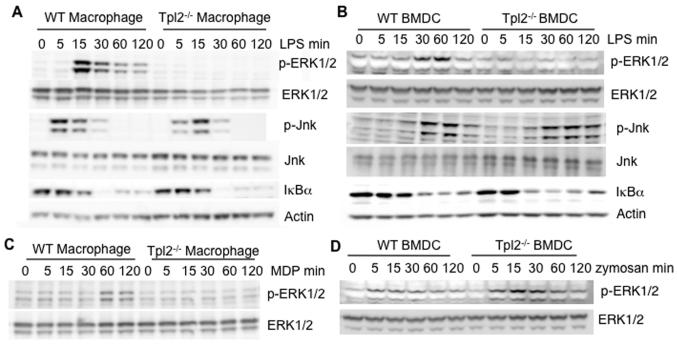
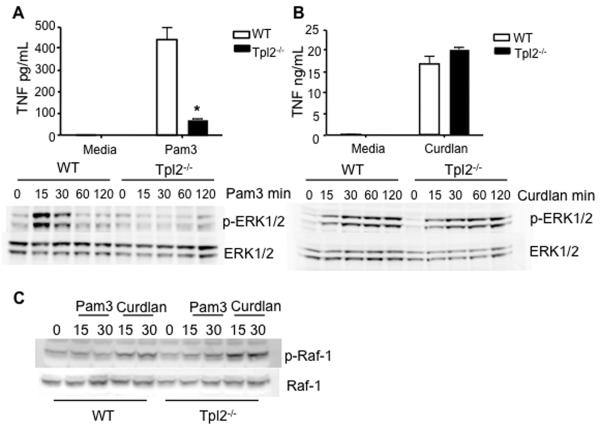
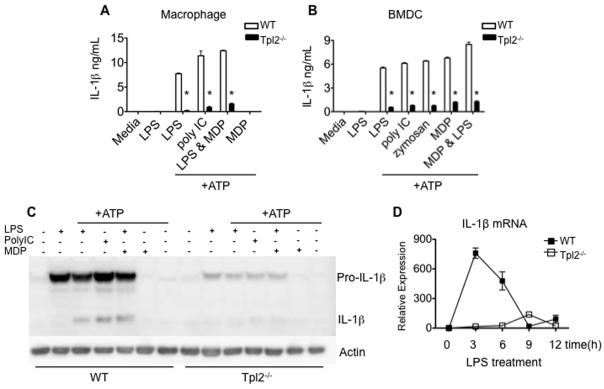
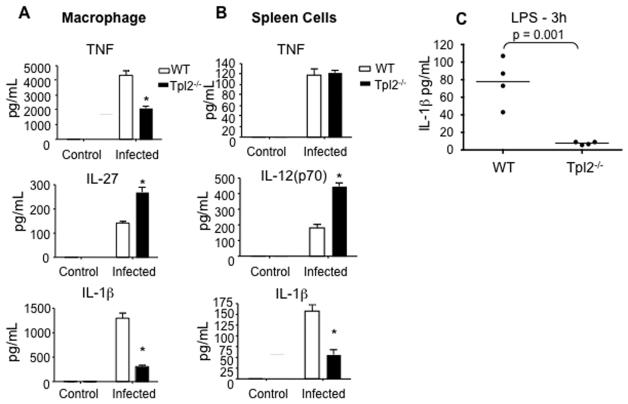
Tumor progression locus 2 (Tpl2, also known as Map3k8 and Cot) is a serine-threonine kinase critical in innate immunity, linking toll-like receptors (TLRs) to TNF production through its activation of ERK. Tpl2(-/-) macrophages have abrogated TNF production but overproduce IL-12 in response to TLR ligands. Despite enhanced IL-12 production, Tpl2(-/-) T cells have impaired IFN-gamma production. Therefore, the role of Tpl2 in a bona fide bacterial infection where all of these cytokines are important in host defense is unclear. To address this issue, we infected Tpl2(-/-) mice with the model pathogen Listeria monocytogenes. We found that Tpl2(-/-) mice infected i.v. with L. monocytogenes had increased pathogen burdens compared with wild-type mice and rapidly succumbed to infection. Enhanced susceptibility correlated with impaired signaling through TLR2 and nucleotide-binding oligomerization domain 2, two receptors previously shown to mediate Listeria recognition. Surprisingly, TNF production in response to infection was not significantly impaired, even though Tpl2 has been implicated in the regulation of TNF. We found that the role of Tpl2 has cell-type specific effects in regulating TNF and transduces signals from some, but not all, pattern recognition receptors (PRR). In contrast to the cell-type- and receptor-specific regulation of TNF, we found that Tpl2 is essential for IL-1beta production from both macrophages and dendritic cells. These studies implicate Tpl2 as an important mediator for collaboration of pattern recognition receptors with danger-associated molecular patterns to induce TNF and IL-1beta production and optimal host defense.
Figures
References
-
- O'Neill LA. When signaling pathways collide: positive and negative regulation of toll-like receptor signal transduction. Immunity. 2008;29:12–20. - PubMed
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Ogura Y, Inohara N, Benito A, Chen FF, Yamaoka S, Nunez G. Nod2, a Nod1/Apaf-1 family member that is restricted to monocytes and activates NF-kappaB. J Biol Chem. 2001;276:4812–4818. - PubMed
-
- Girardin SE, Boneca IG, Viala J, Chamaillard M, Labigne A, Thomas G, Philpott DJ, Sansonetti PJ. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J Biol Chem. 2003;278:8869–8872. - PubMed
-
- Inohara N, Ogura Y, Fontalba A, Gutierrez O, Pons F, Crespo J, Fukase K, Inamura S, Kusumoto S, Hashimoto M, Foster SJ, Moran AP, Fernandez-Luna JL, Nunez G. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn's disease. J Biol Chem. 2003;278:5509–5512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1 K22 AR53953-01/AR/NIAMS NIH HHS/United States
- ZIA AR041161/ImNIH/Intramural NIH HHS/United States
- ZIA AR041106/ImNIH/Intramural NIH HHS/United States
- ZIA AR041167/ImNIH/Intramural NIH HHS/United States
- Z01 AR041106/ImNIH/Intramural NIH HHS/United States
- Z01 AR041160/ImNIH/Intramural NIH HHS/United States
- R01 CA095431/CA/NCI NIH HHS/United States
- Z01 AR041159/ImNIH/Intramural NIH HHS/United States
- K22 AR053953/AR/NIAMS NIH HHS/United States
- Z01 AR041167/ImNIH/Intramural NIH HHS/United States
- Z01 AR041132/ImNIH/Intramural NIH HHS/United States
- ZIA AR041159/ImNIH/Intramural NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
