

Differential effect of saturated and unsaturated free fatty acids on the generation of monocyte adhesion and chemotactic factors by adipocytes: dissociation of adipocyte hypertrophy from inflammation
- PMID: 19934003
- PMCID: PMC2809975
- DOI: 10.2337/db09-0925
Differential effect of saturated and unsaturated free fatty acids on the generation of monocyte adhesion and chemotactic factors by adipocytes: dissociation of adipocyte hypertrophy from inflammation
Abstract
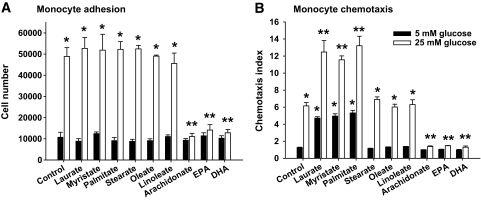
Objective: Obesity is associated with monocyte-macrophage accumulation in adipose tissue. Previously, we showed that glucose-stimulated production by adipocytes of serum amyloid A (SAA), monocyte chemoattractant protein (MCP)-1, and hyaluronan (HA) facilitated monocyte accumulation. The current objective was to determine how the other major nutrient, free fatty acids (FFAs), affects these molecules and monocyte recruitment by adipocytes.
Research design and methods: Differentiated 3T3-L1, Simpson-Golabi-Behmel syndrome adipocytes, and mouse embryonic fibroblasts were exposed to various FFAs (250 micromol/l) in either 5 or 25 mmol/l (high) glucose for evaluation of SAA, MCP-1, and HA regulation in vitro.
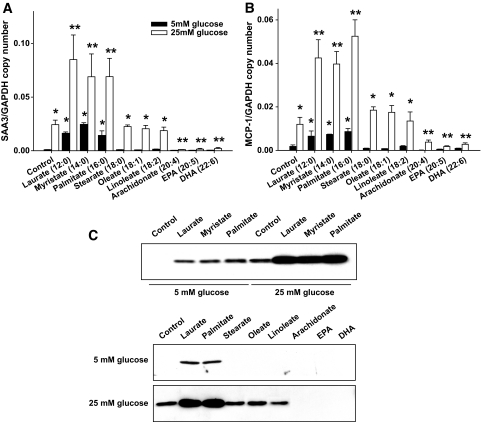
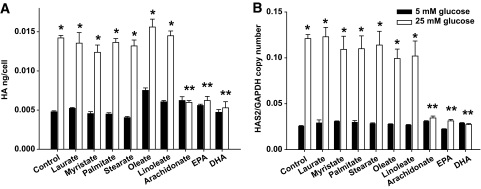
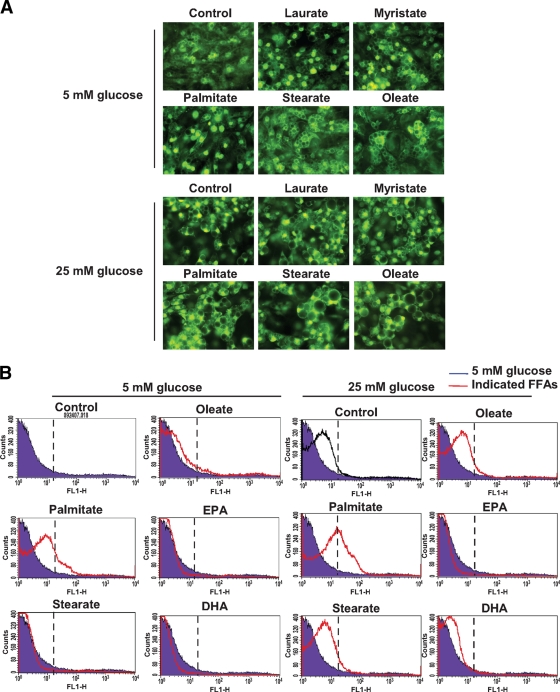
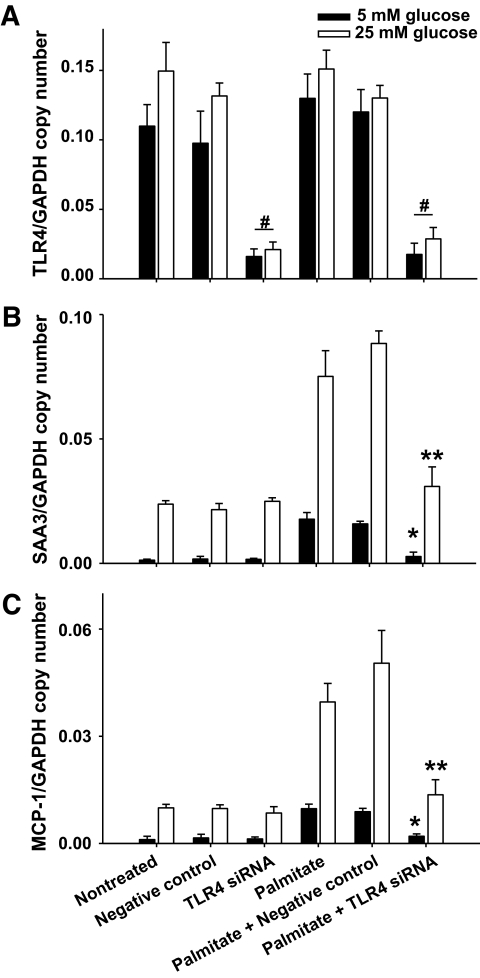
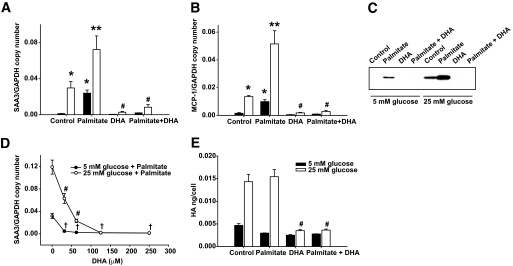
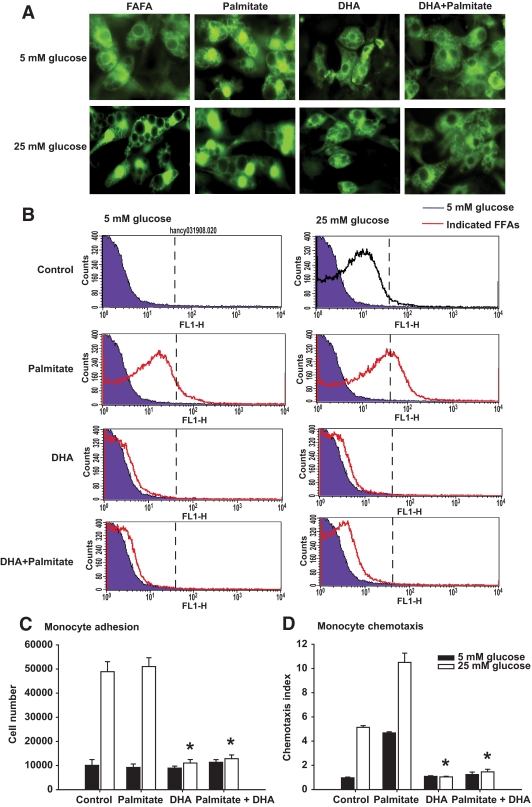
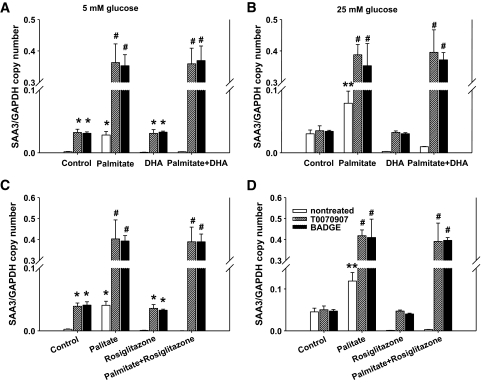
Results: Saturated fatty acids (SFAs) such as laurate, myristate, and palmitate increased cellular triglyceride accumulation, SAA, and MCP-1 expression; generated reactive oxygen species (ROS); and increased nuclear factor (NF) kappaB translocation in both 5 and 25 mmol/l glucose. Conversely, polyunsaturated fatty acids (PUFAs) such as arachidonate, eicosapentaenate, and docosahexaenate (DHA) decreased these events. Gene expression could be dissociated from triglyceride accumulation. Although excess glucose increased HA content, SFAs, oleate, and linoleate did not. Antioxidant treatment repressed glucose- and palmitate-stimulated ROS generation and NFkappaB translocation and decreased SAA and MCP-1 expression and monocyte chemotaxis. Silencing toll-like receptor-4 (TLR4) markedly reduced SAA and MCP-1 expression in response to palmitate but not glucose. DHA suppressed NFkappaB translocation stimulated by both excess glucose and palmitate via a peroxisome prolifterator-activated receptor (PPAR) gamma-dependent pathway.
Conclusions: Excess glucose and SFAs regulate chemotactic factor expression by a mechanism that involves ROS generation, NFkappaB, and PPARgamma, and which is repressed by PUFAs. Certain SFAs, but not excess glucose, trigger chemotactic factor expression via a TLR4-dependent pathway.
Figures
References
-
- Bullo M, Garcia-Lorda P, Megias I, Salas-Salvado J: Systemic inflammation, adipose tissue tumor necrosis factor, and leptin expression. Obes Res 2003;11:525–531 - PubMed
-
- Maachi M, Pieroni L, Bruckert E, Jardel C, Fellahi S, Hainque B, Capeau J, Bastard JP: Systemic low-grade inflammation is related to both circulating and adipose tissue TNFalpha, leptin and IL-6 levels in obese women. Int J Obes Relat Metab Disord 2004;28:993–997 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
