

Control in time and space: Tramtrack69 cooperates with Notch and Ecdysone to repress ectopic fate and shape changes during Drosophila egg chamber maturation
- PMID: 19934014
- PMCID: PMC2781054
- DOI: 10.1242/dev.042770
Control in time and space: Tramtrack69 cooperates with Notch and Ecdysone to repress ectopic fate and shape changes during Drosophila egg chamber maturation
Abstract
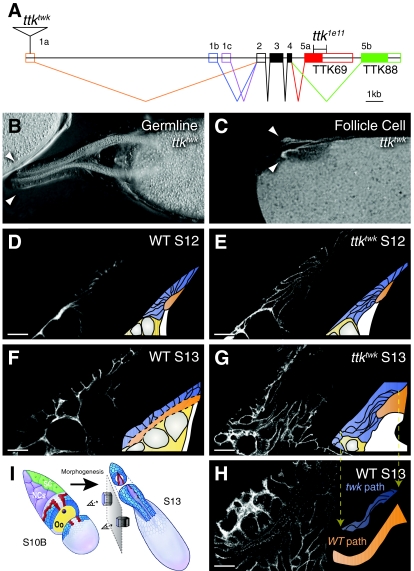
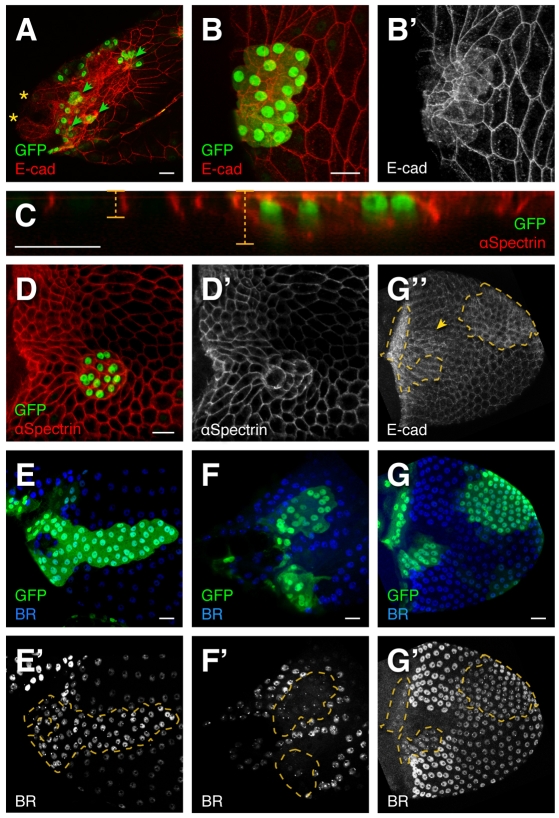
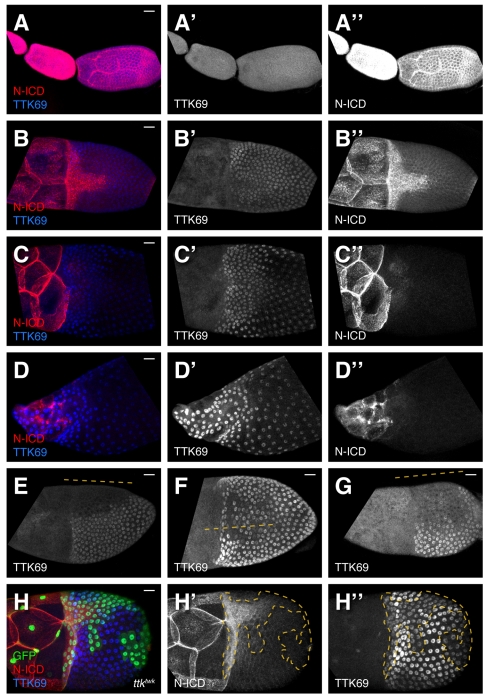
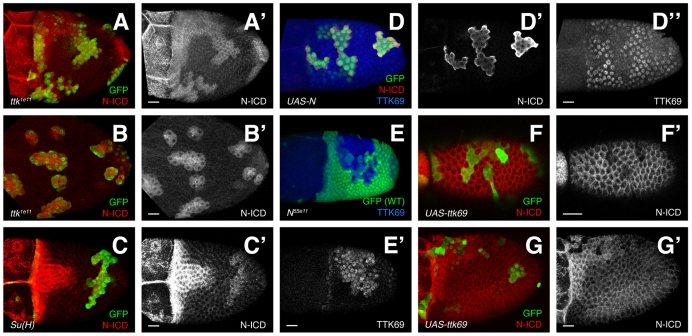
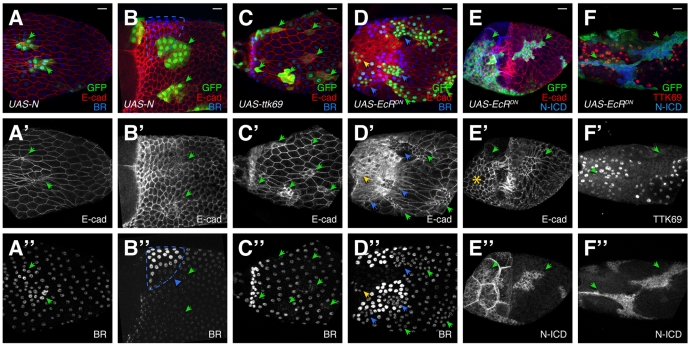
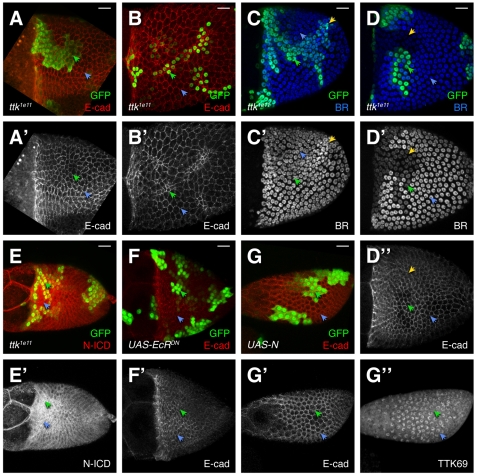
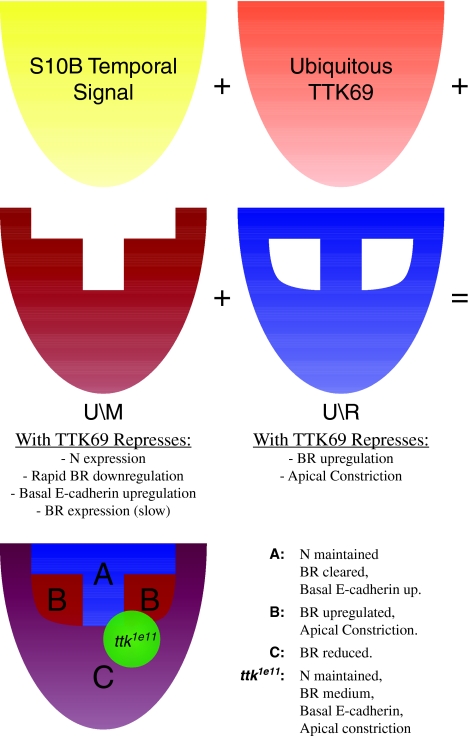
Organ morphogenesis requires cooperation between cells, which determine their course of action based upon location within a tissue. Just as important, cells must synchronize their activities, which requires awareness of developmental time. To understand how cells coordinate behaviors in time and space, we analyzed Drosophila egg chamber development. We found that the transcription factor Tramtrack69 (TTK69) controls the fates and shapes of all columnar follicle cells by integrating temporal and spatial information, restricting characteristic changes in morphology and expression that occur at stage 10B to appropriate domains. TTK69 is required again later in oogenesis: it controls the volume of the dorsal-appendage (DA) tubes by promoting apical re-expansion and lateral shortening of DA-forming follicle cells. We show that TTK69 and Notch compete to repress each other's expression and that a local Ecdysone signal is required to shift the balance in favor of TTK69. We hypothesize that TTK69 then cooperates with spatially restricted co-factors to define appropriate responses to a globally available (but as yet unidentified) temporal signal that initiates the S10B transformations.
Figures
References
-
- Araújo S. J., Cela C., Llimargas M. (2007). Tramtrack regulates different morphogenetic events during Drosophila tracheal development. Development 134, 3665-3676 - PubMed
-
- Ashe H. L., Briscoe J. (2006). The interpretation of morphogen gradients. Development 133, 385-394 - PubMed
-
- Badenhorst P. (2001). Tramtrack controls glial number and identity in the Drosophila embryonic CNS. Development 128, 4093-4101 - PubMed
-
- Banerjee I., Clayton P. (2007). The genetic basis for the timing of human puberty. J. Neuroendocrinol. 19, 831-838 - PubMed
-
- Baonza A., Murawsky C. M., Travers A. A., Freeman M. (2002). Pointed and Tramtrack69 establish an EGFR-dependent transcriptional switch to regulate mitosis. Nat. Cell Biol. 4, 976-980 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
