

Novel oligoamine analogues inhibit lysine-specific demethylase 1 and induce reexpression of epigenetically silenced genes
- PMID: 19934284
- PMCID: PMC2927136
- DOI: 10.1158/1078-0432.CCR-09-1293
Novel oligoamine analogues inhibit lysine-specific demethylase 1 and induce reexpression of epigenetically silenced genes
Abstract
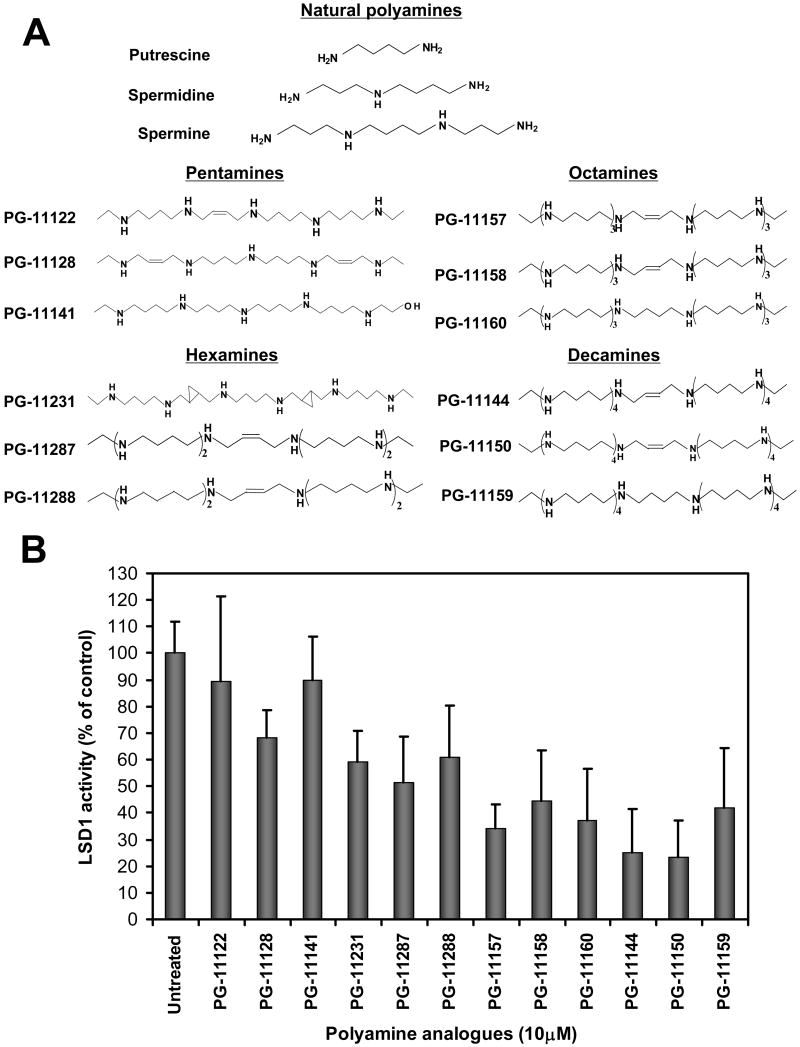
Purpose: Abnormal DNA CpG island hypermethylation and transcriptionally repressive histone modifications are associated with the aberrant silencing of tumor suppressor genes. Lysine methylation is a dynamic, enzymatically controlled process. Lysine-specific demethylase 1 (LSD1) has recently been identified as a histone lysine demethylase. LSD1 specifically catalyzes demethylation of mono- and dimethyl-lysine 4 of histone 3 (H3K4), key positive chromatin marks associated with transcriptional activation. We hypothesized that a novel class of oligoamine analogues would effectively inhibit LSD1 and thus cause the reexpression of aberrantly silenced genes.
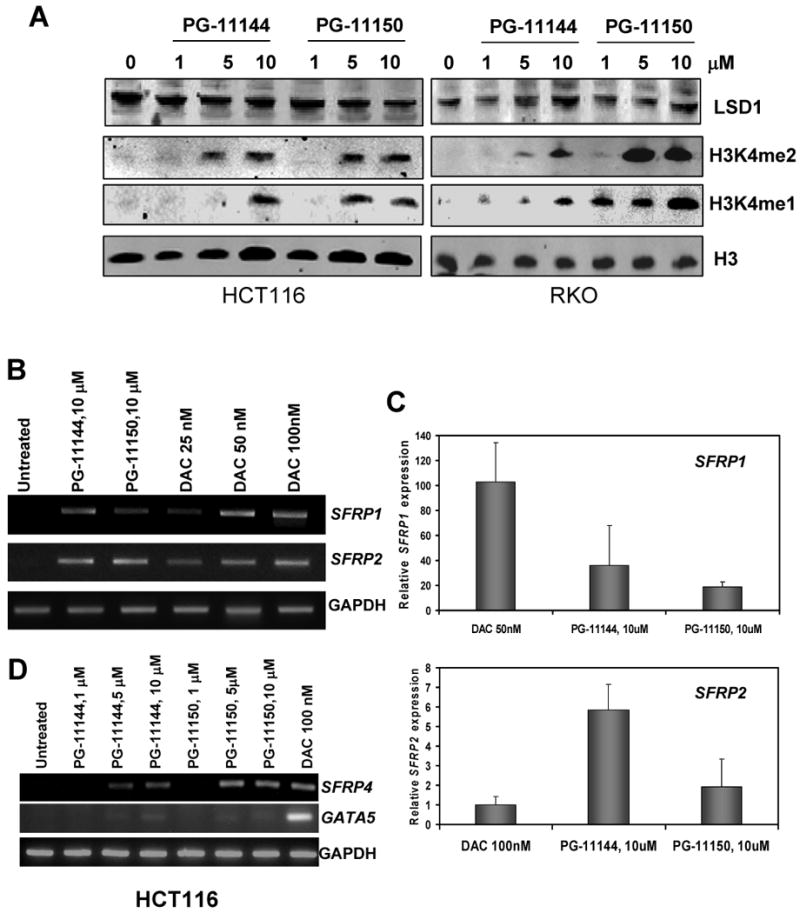
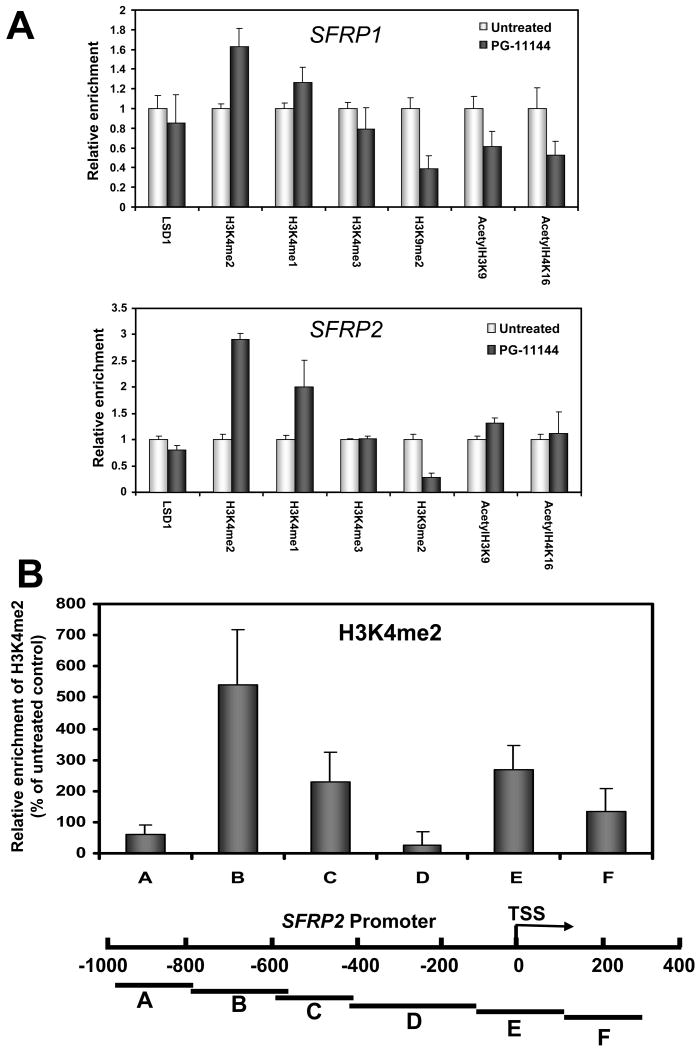
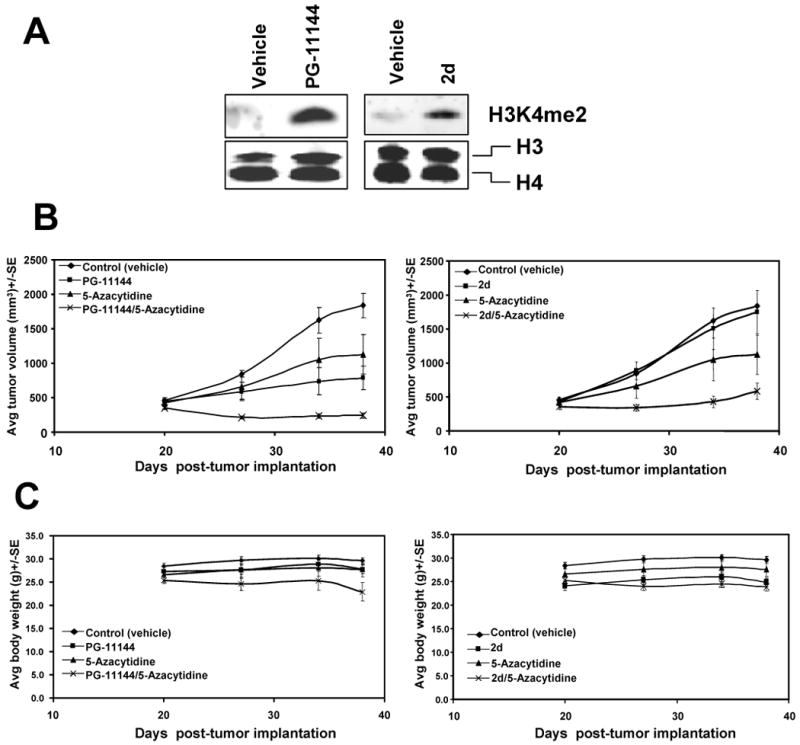
Experimental design: Human colorectal cancer cells were treated with the oligoamines and changes in mono- and dimethyl-H3K4 and other chromatin marks were monitored. In addition, treated cells were evaluated for the reexpression of the aberrantly silenced secreted frizzled-related proteins (SFRP) Wnt signaling pathway antagonist genes. Finally, the effects of the LSD1 inhibitors were evaluated in an in vivo xenograft model.
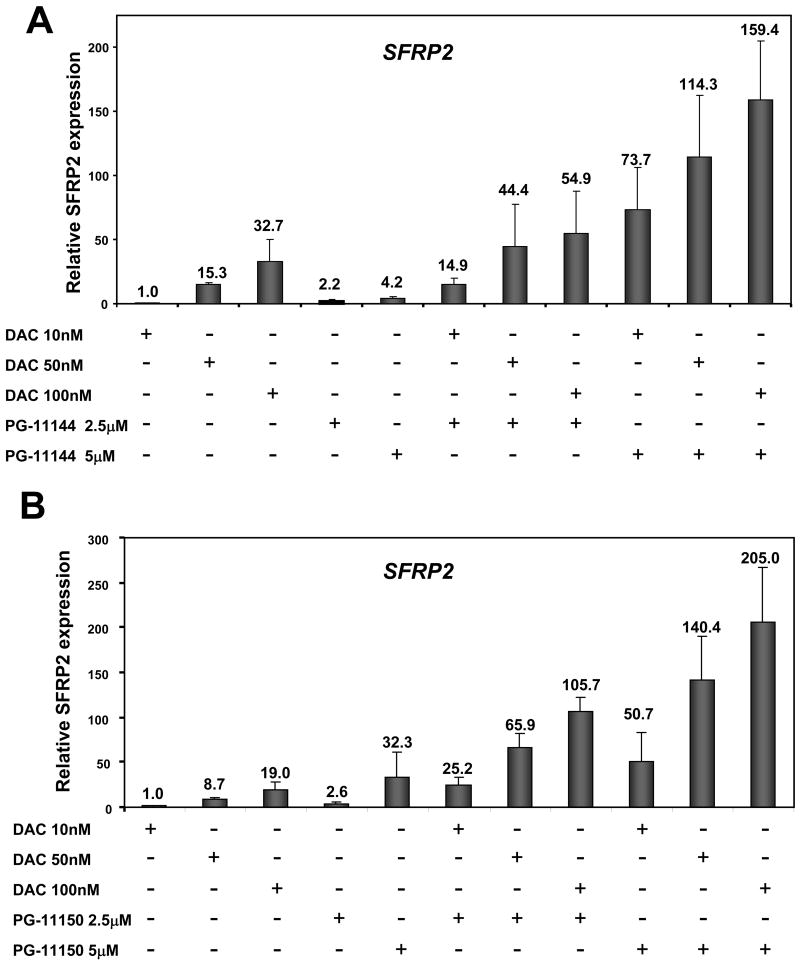
Results: Treatment of HCT116 human colon adenocarcinoma cells in vitro resulted in increased H3K4 methylation and reexpression of silenced SFRP genes. This reexpression is also accompanied by a decrease in H3K9me2 repressive mark. Importantly, cotreatment with low doses of oligoamines and a DNA methyltransferase inhibitor highly induces the reexpression of the aberrantly silenced SFRP2 gene and results in significant inhibition of the growth of established tumors in a human colon tumor model in vivo.
Conclusions: The use of LSD1-inhibiting oligoamine analogues in combination with DNA methyltransferase inhibitors represents a highly promising and novel approach for epigenetic therapy of cancer.
Conflict of interest statement
Figures

Comment in
-
Targeting histone demethylases in cancer therapy.Clin Cancer Res. 2009 Dec 1;15(23):7111-3. doi: 10.1158/1078-0432.CCR-09-2399. Epub 2009 Nov 24. Clin Cancer Res. 2009. PMID: 19934292 Free PMC article.
References
-
- Baylin SB, Ohm JE. Epigenetic gene silencing in cancer - a mechanism for early oncogenic pathway addiction? Nat Rev Cancer. 2006;6:107–16. - PubMed
-
- Jenuwein T, Allis C. Translating the histone code. Science. 2001;293:1074–80. - PubMed
-
- Shi Y, Lan F, Matson C, et al. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell. 2004;119:941–53. - PubMed
-
- Lee M, Wynder C, Cooch N, Shiekhattar R. An essential role for CoREST in nucleosomal histone 3 lysine 4 demethylation. Nature. 2005;437:432–5. - PubMed
-
- Tsukada Y, Fang J, Erdjument-Bromage H, Warren M, Borchers C, Tempst P, Zhang Y. Histone demethylation by a family of JmjC domain-containing proteins. Nature. 2006;439:811–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
