

Regulators of the cytoplasmic dynein motor
- PMID: 19935668
- PMCID: PMC3394690
- DOI: 10.1038/nrm2804
Regulators of the cytoplasmic dynein motor
Abstract
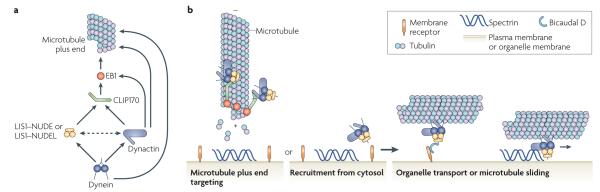
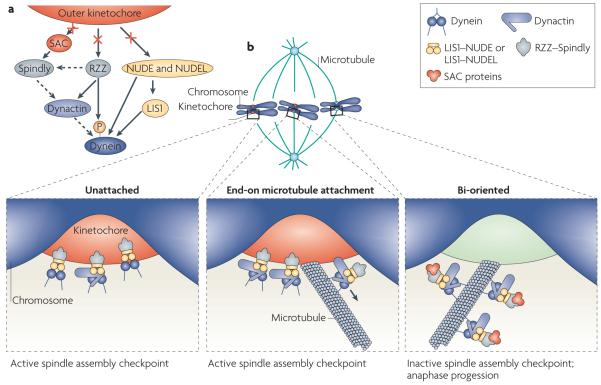
Eukaryotic cells use cytoskeletal motor proteins to transport many different intracellular cargos. Numerous kinesins and myosins have evolved to cope with the various transport needs that have arisen during eukaryotic evolution. Surprisingly, a single cytoplasmic dynein (a minus end-directed microtubule motor) carries out similarly diverse transport activities as the many different types of kinesin. How is dynein coupled to its wide range of cargos and how is it spatially and temporally regulated? The answer could lie in the several multifunctional adaptors, including dynactin, lissencephaly 1, nuclear distribution protein E (NUDE) and NUDE-like, Bicaudal D, Rod-ZW10-Zwilch and Spindly, that regulate dynein function and localization.
Figures
References
-
- Karki S, Holzbaur EL. Cytoplasmic dynein and dynactin in cell division and intracellular transport. Curr. Opin. Cell Biol. 1999;11:45–53. - PubMed
-
- Vale RD. The molecular motor toolbox for intracellular transport. Cell. 2003;112:467–480. - PubMed
-
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. AAA+: a class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999;9:27–43. - PubMed
-
- Karki S, Holzbaur EL. Affinity chromatography demonstrates a direct binding between cytoplasmic dynein and the dynactin complex. J. Biol. Chem. 1995;270:28806–28811. - PubMed
