

A dominant mutation in mediator of paramutation2, one of three second-largest subunits of a plant-specific RNA polymerase, disrupts multiple siRNA silencing processes
- PMID: 19936058
- PMCID: PMC2774164
- DOI: 10.1371/journal.pgen.1000725
A dominant mutation in mediator of paramutation2, one of three second-largest subunits of a plant-specific RNA polymerase, disrupts multiple siRNA silencing processes
Abstract
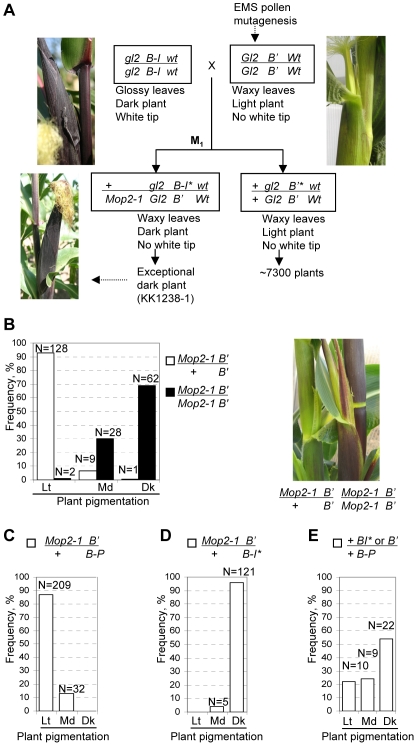
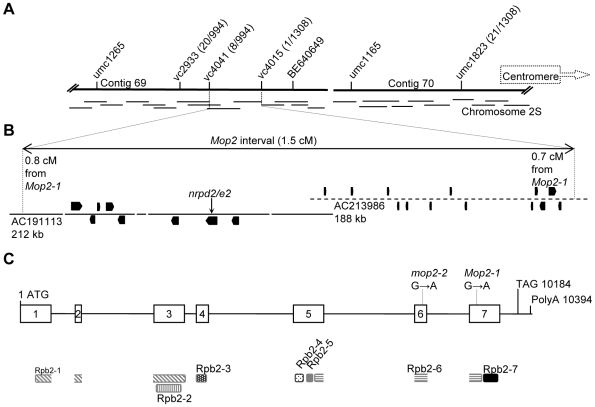
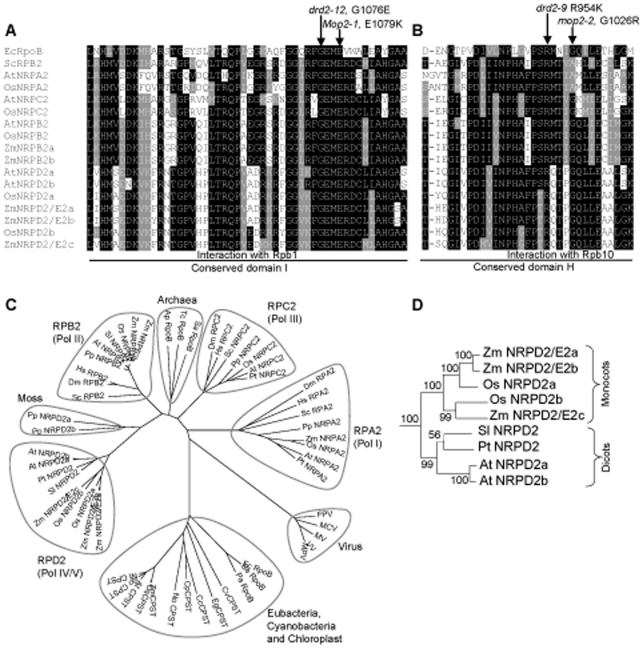
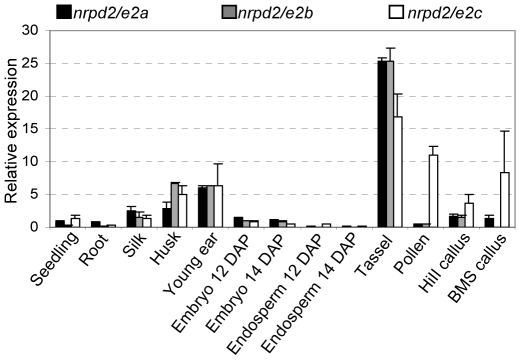
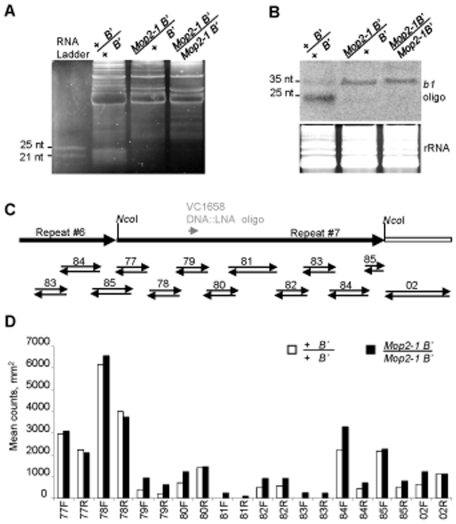
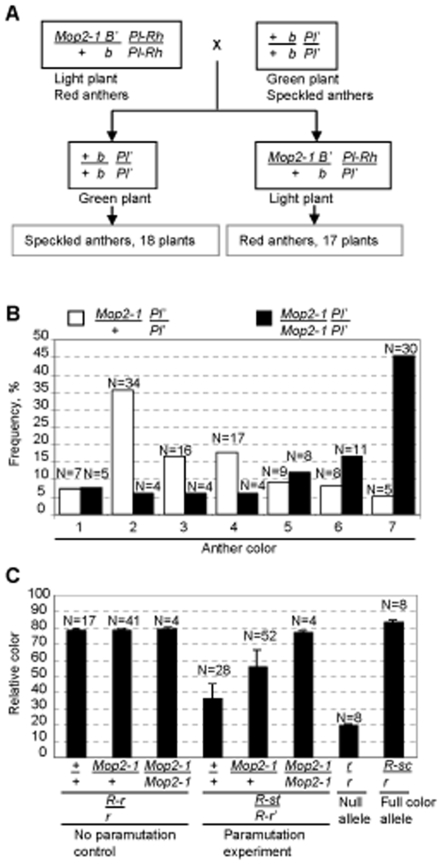
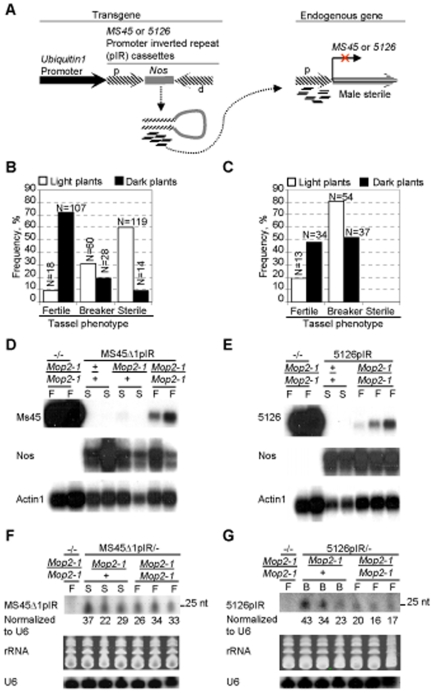
Paramutation involves homologous sequence communication that leads to meiotically heritable transcriptional silencing. We demonstrate that mop2 (mediator of paramutation2), which alters paramutation at multiple loci, encodes a gene similar to Arabidopsis NRPD2/E2, the second-largest subunit of plant-specific RNA polymerases IV and V. In Arabidopsis, Pol-IV and Pol-V play major roles in RNA-mediated silencing and a single second-largest subunit is shared between Pol-IV and Pol-V. Maize encodes three second-largest subunit genes: all three genes potentially encode full length proteins with highly conserved polymerase domains, and each are expressed in multiple overlapping tissues. The isolation of a recessive paramutation mutation in mop2 from a forward genetic screen suggests limited or no functional redundancy of these three genes. Potential alternative Pol-IV/Pol-V-like complexes could provide maize with a greater diversification of RNA-mediated transcriptional silencing machinery relative to Arabidopsis. Mop2-1 disrupts paramutation at multiple loci when heterozygous, whereas previously silenced alleles are only up-regulated when Mop2-1 is homozygous. The dramatic reduction in b1 tandem repeat siRNAs, but no disruption of silencing in Mop2-1 heterozygotes, suggests the major role for tandem repeat siRNAs is not to maintain silencing. Instead, we hypothesize the tandem repeat siRNAs mediate the establishment of the heritable silent state-a process fully disrupted in Mop2-1 heterozygotes. The dominant Mop2-1 mutation, which has a single nucleotide change in a domain highly conserved among all polymerases (E. coli to eukaryotes), disrupts both siRNA biogenesis (Pol-IV-like) and potentially processes downstream (Pol-V-like). These results suggest either the wild-type protein is a subunit in both complexes or the dominant mutant protein disrupts both complexes. Dominant mutations in the same domain in E. coli RNA polymerase suggest a model for Mop2-1 dominance: complexes containing Mop2-1 subunits are non-functional and compete with wild-type complexes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Diverse gene-silencing mechanisms with distinct requirements for RNA polymerase subunits in Zea mays.Genetics. 2014 Nov;198(3):1031-42. doi: 10.1534/genetics.114.168518. Epub 2014 Aug 27. Genetics. 2014. PMID: 25164883 Free PMC article.
-
RNA polymerase IV functions in paramutation in Zea mays.Science. 2009 Feb 27;323(5918):1201-5. doi: 10.1126/science.1164508. Science. 2009. PMID: 19251626
-
RNA-mediated trans-communication can establish paramutation at the b1 locus in maize.Proc Natl Acad Sci U S A. 2010 Jul 20;107(29):12986-91. doi: 10.1073/pnas.1007972107. Epub 2010 Jun 29. Proc Natl Acad Sci U S A. 2010. PMID: 20616013 Free PMC article.
-
Paramutation in maize: RNA mediated trans-generational gene silencing.Curr Opin Genet Dev. 2010 Apr;20(2):156-63. doi: 10.1016/j.gde.2010.01.008. Epub 2010 Feb 12. Curr Opin Genet Dev. 2010. PMID: 20153628 Free PMC article. Review.
-
Paramutation: a process for acquiring trans-generational regulatory states.Curr Opin Plant Biol. 2011 Apr;14(2):210-6. doi: 10.1016/j.pbi.2011.02.005. Epub 2011 Mar 17. Curr Opin Plant Biol. 2011. PMID: 21420347 Review.
Cited by
-
Genome-wide loss of CHH methylation with limited transcriptome changes in Setaria viridis DOMAINS REARRANGED METHYLTRANSFERASE (DRM) mutants.Plant J. 2022 Jul;111(1):103-116. doi: 10.1111/tpj.15781. Epub 2022 May 6. Plant J. 2022. PMID: 35436373 Free PMC article.
-
Non-coding RNA polymerases that silence transposable elements and reprogram gene expression in plants.Transcription. 2020 Jun-Aug;11(3-4):172-191. doi: 10.1080/21541264.2020.1825906. Epub 2020 Nov 12. Transcription. 2020. PMID: 33180661 Free PMC article. Review.
-
Altered nucleosome positions in maize haplotypes and mutants of a subset of SWI/SNF-like proteins.Plant Direct. 2017 Oct 16;1(4):e00019. doi: 10.1002/pld3.19. eCollection 2017 Oct. Plant Direct. 2017. PMID: 31245667 Free PMC article.
-
siRNA-mediated DNA methylation and H3K9 dimethylation in plants.Protein Cell. 2013 Sep;4(9):656-63. doi: 10.1007/s13238-013-3052-7. Epub 2013 Aug 13. Protein Cell. 2013. PMID: 23943321 Free PMC article. Review.
-
RNA-directed DNA methylation enforces boundaries between heterochromatin and euchromatin in the maize genome.Proc Natl Acad Sci U S A. 2015 Nov 24;112(47):14728-33. doi: 10.1073/pnas.1514680112. Epub 2015 Nov 9. Proc Natl Acad Sci U S A. 2015. PMID: 26553984 Free PMC article.
References
-
- Pilu R, Panzeri D, Cassani E, Cerino Badone F, Landoni M, et al. A paramutation phenomenon is involved in the genetics of maize low phytic acid1-241 (lpa1-241) trait. Heredity. 2009;102:236–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
