

Catastrophic NAD+ depletion in activated T lymphocytes through Nampt inhibition reduces demyelination and disability in EAE
- PMID: 19936064
- PMCID: PMC2774509
- DOI: 10.1371/journal.pone.0007897
Catastrophic NAD+ depletion in activated T lymphocytes through Nampt inhibition reduces demyelination and disability in EAE
Abstract
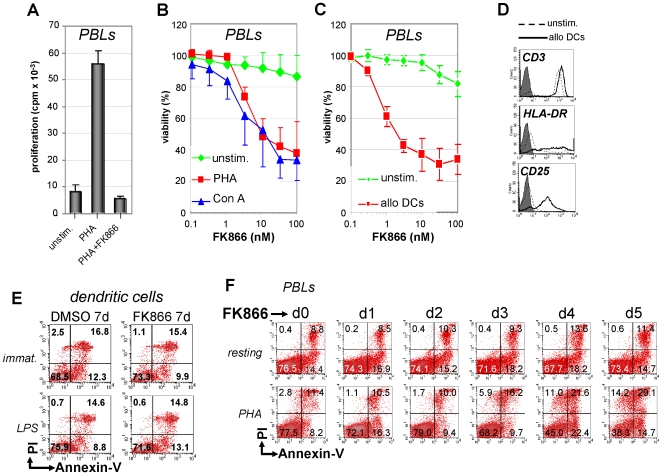
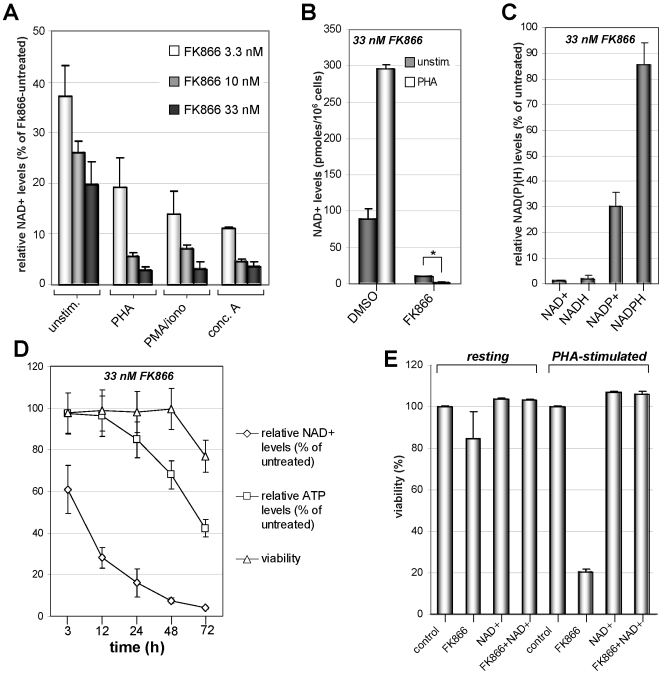
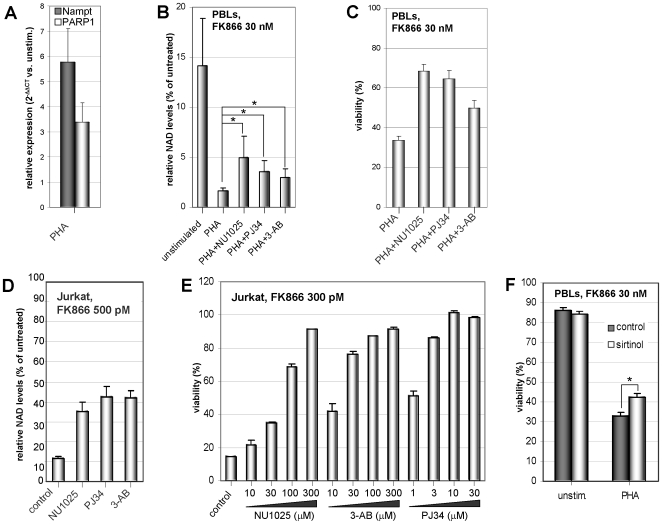
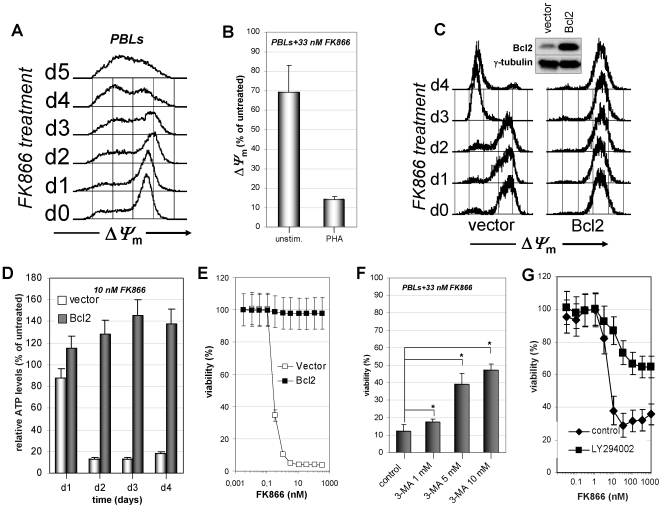
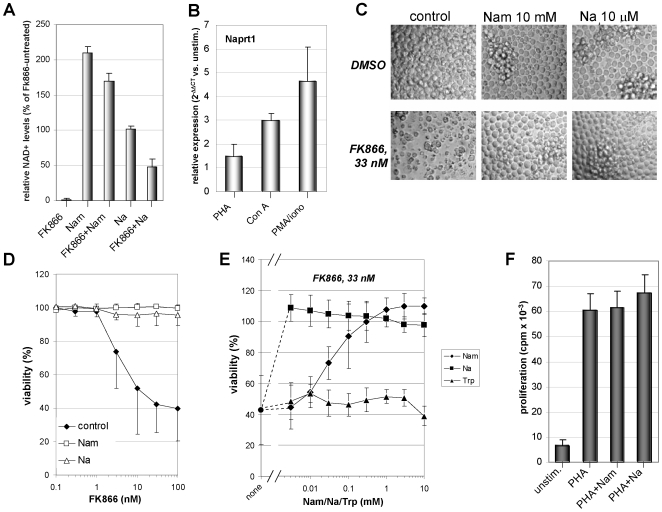
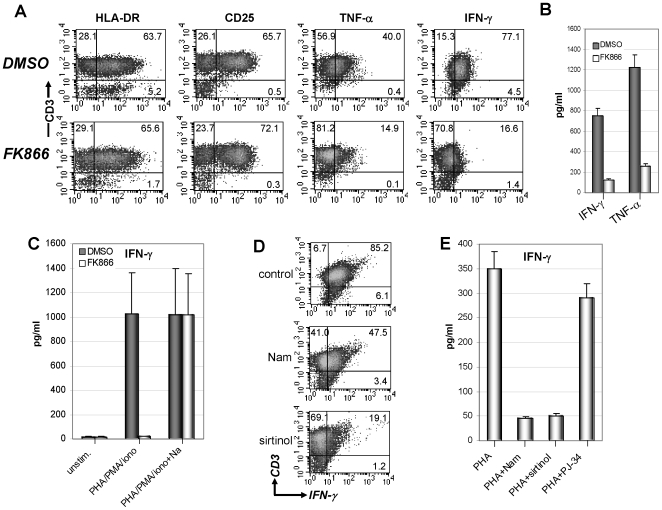
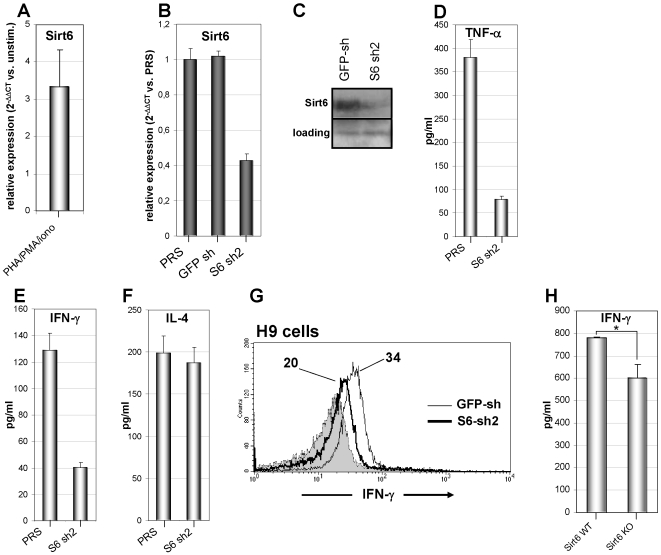
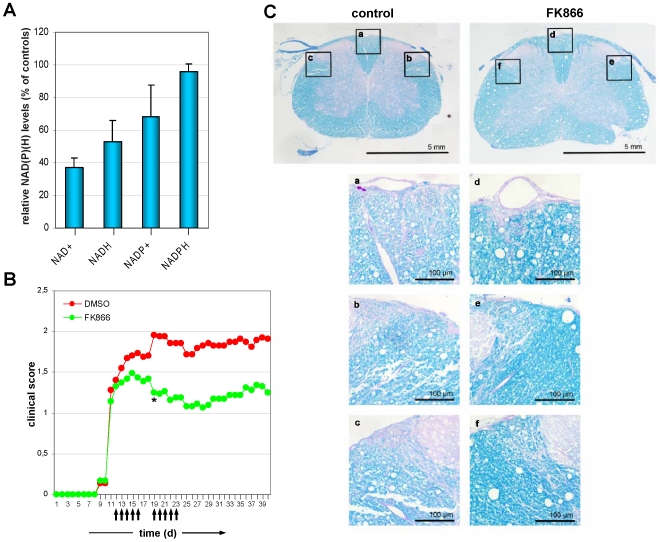
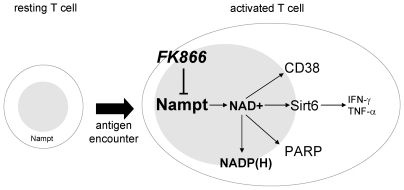
Nicotinamide phosphoribosyltransferase (Nampt) inhibitors such as FK866 are potent inhibitors of NAD(+) synthesis that show promise for the treatment of different forms of cancer. Based on Nampt upregulation in activated T lymphocytes and on preliminary reports of lymphopenia in FK866 treated patients, we have investigated FK866 for its capacity to interfere with T lymphocyte function and survival. Intracellular pyridine nucleotides, ATP, mitochondrial function, viability, proliferation, activation markers and cytokine secretion were assessed in resting and in activated human T lymphocytes. In addition, we used experimental autoimmune encephalomyelitis (EAE) as a model of T-cell mediated autoimmune disease to assess FK866 efficacy in vivo. We show that activated, but not resting, T lymphocytes undergo massive NAD(+) depletion upon FK866-mediated Nampt inhibition. As a consequence, impaired proliferation, reduced IFN-gamma and TNF-alpha production, and finally autophagic cell demise result. We demonstrate that upregulation of the NAD(+)-degrading enzyme poly-(ADP-ribose)-polymerase (PARP) by activated T cells enhances their susceptibility to NAD(+) depletion. In addition, we relate defective IFN-gamma and TNF-alpha production in response to FK866 to impaired Sirt6 activity. Finally, we show that FK866 strikingly reduces the neurological damage and the clinical manifestations of EAE. In conclusion, Nampt inhibitors (and possibly Sirt6 inhibitors) could be used to modulate T cell-mediated immune responses and thereby be beneficial in immune-mediated disorders.
Conflict of interest statement
Figures
Similar articles
-
Nicotinamide phosphoribosyltransferase (NAMPT) activity is essential for survival of resting lymphocytes.Immunol Cell Biol. 2014 Feb;92(2):191-9. doi: 10.1038/icb.2013.85. Epub 2013 Nov 26. Immunol Cell Biol. 2014. PMID: 24275857
-
FK866-induced NAMPT inhibition activates AMPK and downregulates mTOR signaling in hepatocarcinoma cells.Biochem Biophys Res Commun. 2015 Mar 6;458(2):334-40. doi: 10.1016/j.bbrc.2015.01.111. Epub 2015 Feb 3. Biochem Biophys Res Commun. 2015. PMID: 25656579
-
Inhibition of nicotinamide phosphoribosyltransferase: cellular bioenergetics reveals a mitochondrial insensitive NAD pool.J Biol Chem. 2010 Oct 29;285(44):34106-14. doi: 10.1074/jbc.M110.136739. Epub 2010 Aug 19. J Biol Chem. 2010. PMID: 20724478 Free PMC article.
-
Nampt/PBEF/visfatin and cancer.Cancer Biol Ther. 2010 Jul 15;10(2):119-25. doi: 10.4161/cbt.10.2.12581. Epub 2010 Jul 3. Cancer Biol Ther. 2010. PMID: 20647743 Review.
-
Non-canonical roles of NAMPT and PARP in inflammation.Dev Comp Immunol. 2021 Feb;115:103881. doi: 10.1016/j.dci.2020.103881. Epub 2020 Oct 7. Dev Comp Immunol. 2021. PMID: 33038343 Review.
Cited by
-
Physiological and pathophysiological roles of NAMPT and NAD metabolism.Nat Rev Endocrinol. 2015 Sep;11(9):535-46. doi: 10.1038/nrendo.2015.117. Epub 2015 Jul 28. Nat Rev Endocrinol. 2015. PMID: 26215259 Review.
-
SIRT6 regulates TNF-α secretion through hydrolysis of long-chain fatty acyl lysine.Nature. 2013 Apr 4;496(7443):110-3. doi: 10.1038/nature12038. Nature. 2013. PMID: 23552949 Free PMC article.
-
Autophagy modulation in multiple sclerosis and experimental autoimmune encephalomyelitis.Clin Exp Immunol. 2022 Aug 19;209(2):140-150. doi: 10.1093/cei/uxac017. Clin Exp Immunol. 2022. PMID: 35641229 Free PMC article. Review.
-
Adipocytokine profile, cytokine levels and foxp3 expression in multiple sclerosis: a possible link to susceptibility and clinical course of disease.PLoS One. 2013 Oct 3;8(10):e76555. doi: 10.1371/journal.pone.0076555. eCollection 2013. PLoS One. 2013. PMID: 24098530 Free PMC article.
-
Cancer metabolic reprogramming: importance, main features, and potentials for precise targeted anti-cancer therapies.Cancer Biol Med. 2014 Mar;11(1):1-19. doi: 10.7497/j.issn.2095-3941.2014.01.001. Cancer Biol Med. 2014. PMID: 24738035 Free PMC article. Review.
References
-
- Hasmann M, Schemainda I. FK866, a highly specific noncompetitive inhibitor of nicotinamide phosphoribosyltransferase, represents a novel mechanism for induction of tumor cell apoptosis. Cancer Res. 2003;63:7436–7442. - PubMed
-
- Khan JA, Tao X, Tong L. Molecular basis for the inhibition of human NMPRTase, a novel target for anticancer agents. Nat Struct Mol Biol. 2006;13:582–588. - PubMed
-
- Wosikowski K, Mattern K, Schemainda I, Hasmann M, Rattel B, et al. WK175, a novel antitumor agent, decreases the intracellular nicotinamide adenine dinucleotide concentration and induces the apoptotic cascade in human leukemia cells. Cancer Res. 2002;62:1057–1062. - PubMed
-
- Nahimana A, Attinger A, Aubry D, Greaney P, Ireson C, et al. The NAD biosynthesis inhibitor APO866 has potent antitumor activity against hematologic malignancies. Blood. 2009;113:3276–3286. - PubMed
-
- Holen K, Saltz LB, Hollywood E, Burk K, Hanauske AR. The pharmacokinetics, toxicities, and biologic effects of FK866, a nicotinamide adenine dinucleotide biosynthesis inhibitor. Invest New Drugs. 2008;26:45–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
