

Modulation of outer hair cell electromotility by cochlear supporting cells and gap junctions
- PMID: 19936276
- PMCID: PMC2775161
- DOI: 10.1371/journal.pone.0007923
Modulation of outer hair cell electromotility by cochlear supporting cells and gap junctions
Abstract
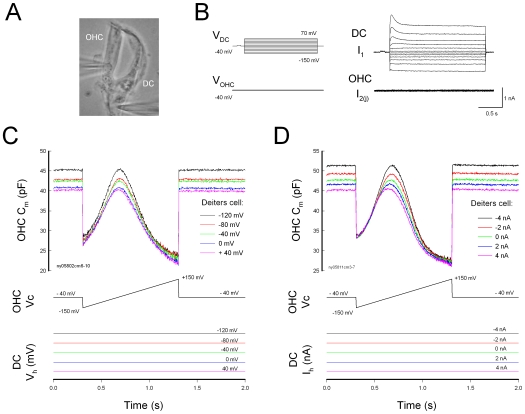
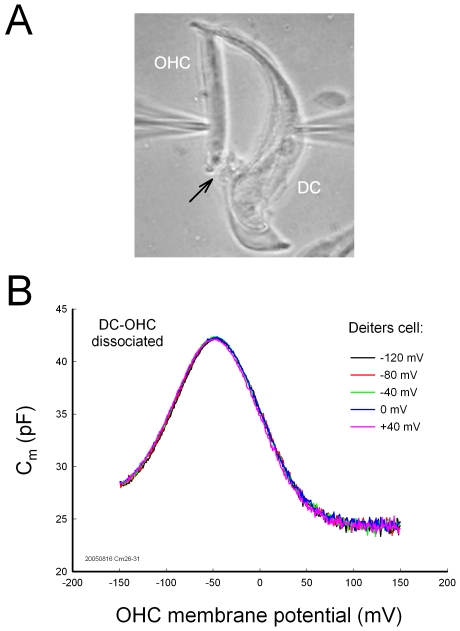
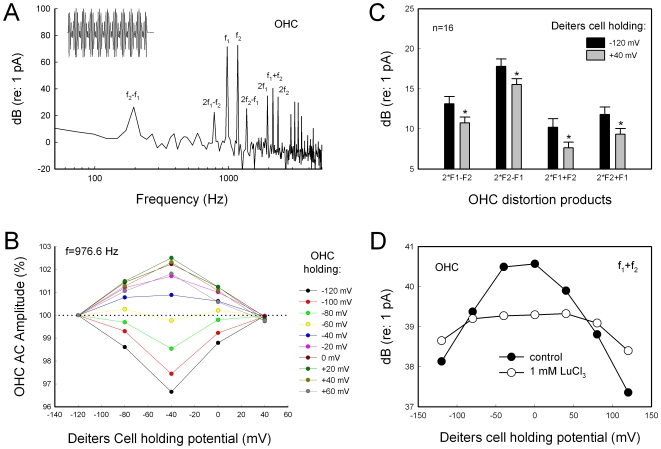
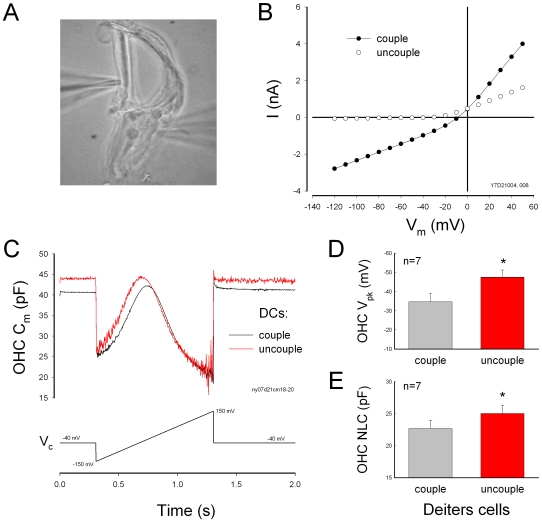
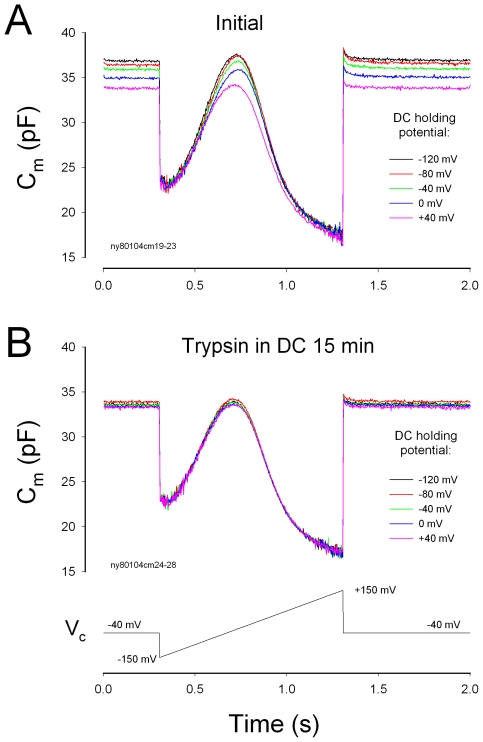
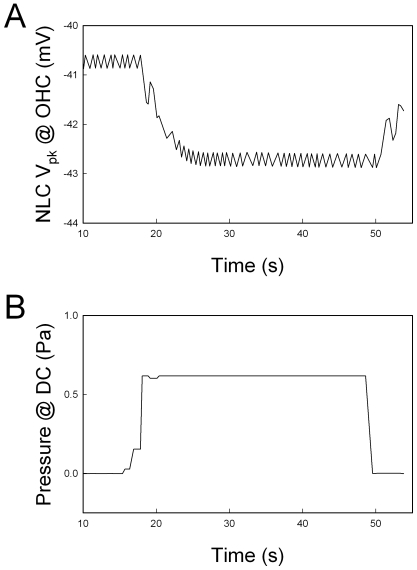
Outer hair cell (OHC) or prestin-based electromotility is an active cochlear amplifier in the mammalian inner ear that can increase hearing sensitivity and frequency selectivity. In situ, Deiters supporting cells are well-coupled by gap junctions and constrain OHCs standing on the basilar membrane. Here, we report that both electrical and mechanical stimulations in Deiters cells (DCs) can modulate OHC electromotility. There was no direct electrical conductance between the DCs and the OHCs. However, depolarization in DCs reduced OHC electromotility associated nonlinear capacitance (NLC) and distortion products. Increase in the turgor pressure of DCs also shifted OHC NLC to the negative voltage direction. Destruction of the cytoskeleton in DCs or dissociation of the mechanical-coupling between DCs and OHCs abolished these effects, indicating the modulation through the cytoskeleton activation and DC-OHC mechanical coupling rather than via electric field potentials. We also found that changes in gap junctional coupling between DCs induced large membrane potential and current changes in the DCs and shifted OHC NLC. Uncoupling of gap junctions between DCs shifted NLC to the negative direction. These data indicate that DCs not only provide a physical scaffold to support OHCs but also can directly modulate OHC electromotility through the DC-OHC mechanical coupling. Our findings reveal a new mechanism of cochlear supporting cells and gap junctional coupling to modulate OHC electromotility and eventually hearing sensitivity in the inner ear.
Conflict of interest statement
Figures
Similar articles
-
Outer hair cell electromotility is low-pass filtered relative to the molecular conformational changes that produce nonlinear capacitance.J Gen Physiol. 2019 Dec 2;151(12):1369-1385. doi: 10.1085/jgp.201812280. Epub 2019 Nov 1. J Gen Physiol. 2019. PMID: 31676485 Free PMC article.
-
A Gap-Junction Mutation Reveals That Outer Hair Cell Extracellular Receptor Potentials Drive High-Frequency Cochlear Amplification.J Neurosci. 2022 Oct 19;42(42):7875-7884. doi: 10.1523/JNEUROSCI.2241-21.2022. Epub 2022 Sep 9. J Neurosci. 2022. PMID: 36261265 Free PMC article.
-
Cochlear outer hair cells in a dominant-negative connexin26 mutant mouse preserve non-linear capacitance in spite of impaired distortion product otoacoustic emission.Neuroscience. 2009 Dec 15;164(3):1312-9. doi: 10.1016/j.neuroscience.2009.08.043. Epub 2009 Aug 25. Neuroscience. 2009. PMID: 19712724
-
Prestin and electromotility may serve multiple roles in cochlear outer hair cells.Hear Res. 2022 Sep 15;423:108428. doi: 10.1016/j.heares.2021.108428. Epub 2021 Dec 26. Hear Res. 2022. PMID: 34987016 Review.
-
The Remarkable Outer Hair Cell: Proceedings of a Symposium in Honour of W. E. Brownell.J Assoc Res Otolaryngol. 2023 Apr;24(2):117-127. doi: 10.1007/s10162-022-00852-4. Epub 2023 Jan 17. J Assoc Res Otolaryngol. 2023. PMID: 36648734 Free PMC article. Review.
Cited by
-
Optogenetics Reveals Roles for Supporting Cells in Force Transmission to and From Outer Hair Cells in the Mouse Cochlea.J Neurosci. 2024 Jan 24;44(4):e1179232023. doi: 10.1523/JNEUROSCI.1179-23.2023. J Neurosci. 2024. PMID: 38050104 Free PMC article.
-
Efferent neurons control hearing sensitivity and protect hearing from noise through the regulation of gap junctions between cochlear supporting cells.J Neurophysiol. 2022 Jan 1;127(1):313-327. doi: 10.1152/jn.00468.2021. Epub 2021 Dec 15. J Neurophysiol. 2022. PMID: 34907797 Free PMC article.
-
Cell degeneration is not a primary causer for Connexin26 (GJB2) deficiency associated hearing loss.Neurosci Lett. 2012 Oct 18;528(1):36-41. doi: 10.1016/j.neulet.2012.08.085. Epub 2012 Sep 11. Neurosci Lett. 2012. PMID: 22975134 Free PMC article.
-
Adenosine Triphosphate (ATP) Inhibits Voltage-Sensitive Potassium Currents in Isolated Hensen's Cells and Nifedipine Protects Against Noise-Induced Hearing Loss in Guinea Pigs.Med Sci Monit. 2016 Jun 13;22:2006-12. doi: 10.12659/msm.898150. Med Sci Monit. 2016. PMID: 27292522 Free PMC article.
-
The endocochlear potential alters cochlear micromechanics.Biophys J. 2011 Jun 8;100(11):2586-94. doi: 10.1016/j.bpj.2011.05.002. Biophys J. 2011. PMID: 21641303 Free PMC article.
References
-
- Forge A, Becker D, Casalotti S, Edwards J, Marziano N, et al. Gap junctions in the inner ear: comparison of distribution patterns in different vertebrates and assessement of connexin composition in mammals. J Comp Neurol. 2003;467:207–231. - PubMed
-
- Slepecky NB. in The cochlea, . In: Dallos P, Popper AN, Fay RR, editors. New York: Springer; 1996. pp. 44–129.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
