

A single amino acid difference in human APOBEC3H variants determines HIV-1 Vif sensitivity
- PMID: 19939923
- PMCID: PMC2812409
- DOI: 10.1128/JVI.01509-09
A single amino acid difference in human APOBEC3H variants determines HIV-1 Vif sensitivity
Abstract
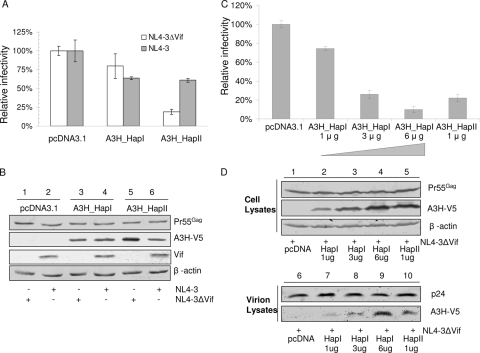
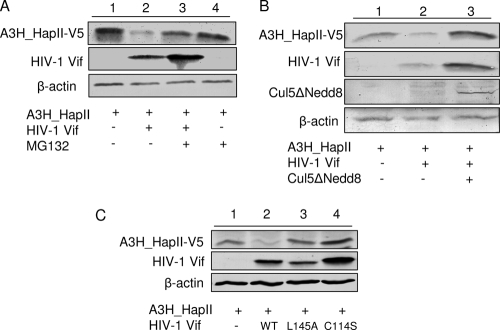
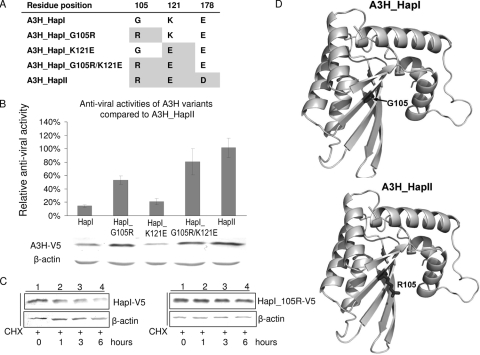
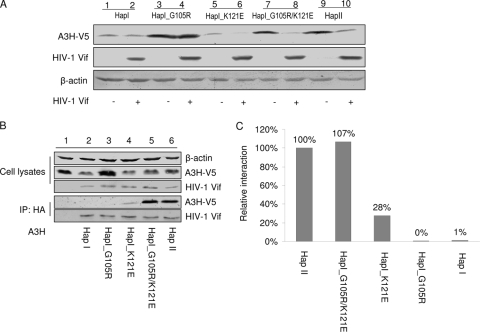
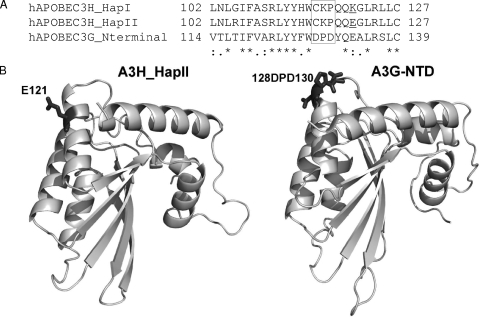
Several variants of APOBEC3H (A3H) have been identified in different human populations. Certain variants of this protein are particularly potent inhibitors of retrotransposons and retroviruses, including HIV-1. However, it is not clear whether HIV-1 Vif can recognize and suppress the antiviral activity of A3H variants, as it does with other APOBEC3 proteins. We now report that A3H_Haplotype II (HapII), a potent inhibitor of HIV-1 in the absence of Vif, can indeed be degraded by HIV-1 Vif. Vif-induced degradation of A3H_HapII was blocked by the proteasome inhibitor MG132 and a Cullin5 (Cul5) dominant negative mutant. In addition, Vif mutants that were incapable of assembly with the host E3 ligase complex factors Cul5, ElonginB, and ElonginC were also defective for A3H_HapII suppression. Although we found that Vif hijacks the same E3 ligase to degrade A3H_HapII as it does to inactivate APOBEC3G (A3G) and APOBEC3F (A3F), more Vif motifs were involved in A3H_HapII inactivation than in either A3G or A3F suppression. In contrast to A3H_HapII, A3H_Haplotype I (HapI), which differs in only three amino acids from A3H_HapII, was resistant to HIV-1 Vif-mediated degradation. We also found that residue 121 was critical for determining A3H sensitivity and binding to HIV-1 Vif.
Figures

Similar articles
-
The activity spectrum of Vif from multiple HIV-1 subtypes against APOBEC3G, APOBEC3F, and APOBEC3H.J Virol. 2012 Jan;86(1):49-59. doi: 10.1128/JVI.06082-11. Epub 2011 Oct 19. J Virol. 2012. PMID: 22013041 Free PMC article.
-
The resistance of human APOBEC3H to HIV-1 NL4-3 molecular clone is determined by a single amino acid in Vif.PLoS One. 2013;8(2):e57744. doi: 10.1371/journal.pone.0057744. Epub 2013 Feb 28. PLoS One. 2013. PMID: 23469063 Free PMC article.
-
APOBEC3G and APOBEC3F Act in Concert To Extinguish HIV-1 Replication.J Virol. 2016 Apr 14;90(9):4681-4695. doi: 10.1128/JVI.03275-15. Print 2016 May. J Virol. 2016. PMID: 26912618 Free PMC article.
-
Degradation-Independent Inhibition of APOBEC3G by the HIV-1 Vif Protein.Viruses. 2021 Apr 3;13(4):617. doi: 10.3390/v13040617. Viruses. 2021. PMID: 33916704 Free PMC article. Review.
-
Host restriction of HIV-1 by APOBEC3 and viral evasion through Vif.Curr Top Microbiol Immunol. 2009;339:1-25. doi: 10.1007/978-3-642-02175-6_1. Curr Top Microbiol Immunol. 2009. PMID: 20012521 Review.
Cited by
-
Human and rhesus APOBEC3D, APOBEC3F, APOBEC3G, and APOBEC3H demonstrate a conserved capacity to restrict Vif-deficient HIV-1.J Virol. 2011 Nov;85(21):11220-34. doi: 10.1128/JVI.05238-11. Epub 2011 Aug 10. J Virol. 2011. PMID: 21835787 Free PMC article.
-
Analysis of the N-terminal positively charged residues of the simian immunodeficiency virus Vif reveals a critical amino acid required for the antagonism of rhesus APOBEC3D, G, and H.Virology. 2014 Jan 20;449:140-9. doi: 10.1016/j.virol.2013.10.037. Epub 2013 Dec 5. Virology. 2014. PMID: 24418547 Free PMC article.
-
A Naturally Occurring Domestic Cat APOBEC3 Variant Confers Resistance to Feline Immunodeficiency Virus Infection.J Virol. 2015 Oct 21;90(1):474-85. doi: 10.1128/JVI.02612-15. Print 2016 Jan 1. J Virol. 2015. PMID: 26491161 Free PMC article.
-
Polymorphism in human APOBEC3H affects a phenotype dominant for subcellular localization and antiviral activity.J Virol. 2011 Aug;85(16):8197-207. doi: 10.1128/JVI.00624-11. Epub 2011 Jun 8. J Virol. 2011. PMID: 21653666 Free PMC article.
-
APOBEC3 degradation is the primary function of HIV-1 Vif determining virion infectivity in the myeloid cell line THP-1.mBio. 2023 Aug 31;14(4):e0078223. doi: 10.1128/mbio.00782-23. Epub 2023 Aug 9. mBio. 2023. PMID: 37555667 Free PMC article.
References
-
- Battey, J. N., J. Kopp, L. Bordoli, R. J. Read, N. D. Clarke, and T. Schwede. 2007. Automated server predictions in CASP7. Proteins 69(Suppl. 8):68-82. - PubMed
-
- Bieniasz, P. D. 2004. Intrinsic immunity: a front-line defense against viral attack. Nat. Immunol. 5:1109-1115. - PubMed
-
- Bishop, K. N., R. K. Holmes, A. M. Sheehy, N. O. Davidson, S. J. Cho, and M. H. Malim. 2004. Cytidine deamination of retroviral DNA by diverse APOBEC proteins. Curr. Biol. 14:1392-1396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
