

Synthesis of heparan sulfate with cyclophilin B-binding properties is determined by cell type-specific expression of sulfotransferases
- PMID: 19940140
- PMCID: PMC2804328
- DOI: 10.1074/jbc.M109.018184
Synthesis of heparan sulfate with cyclophilin B-binding properties is determined by cell type-specific expression of sulfotransferases
Abstract
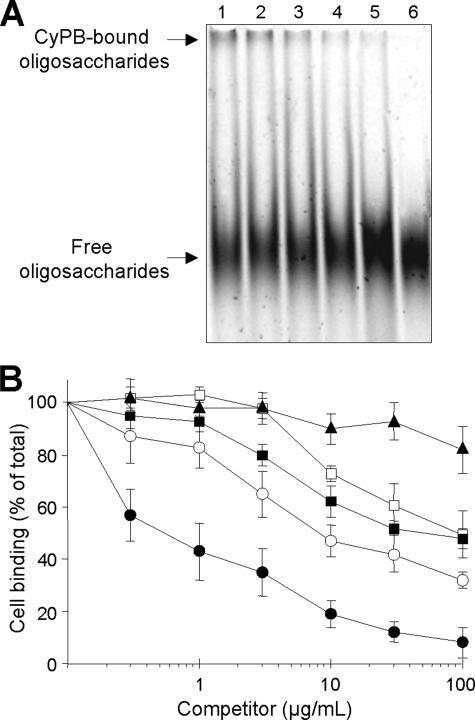
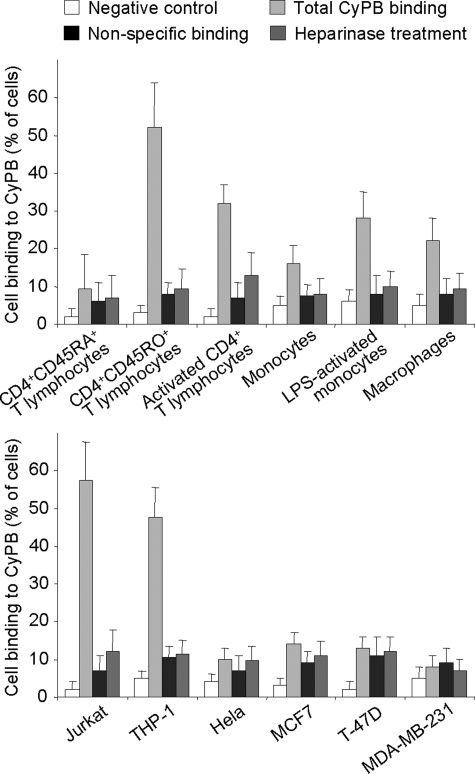
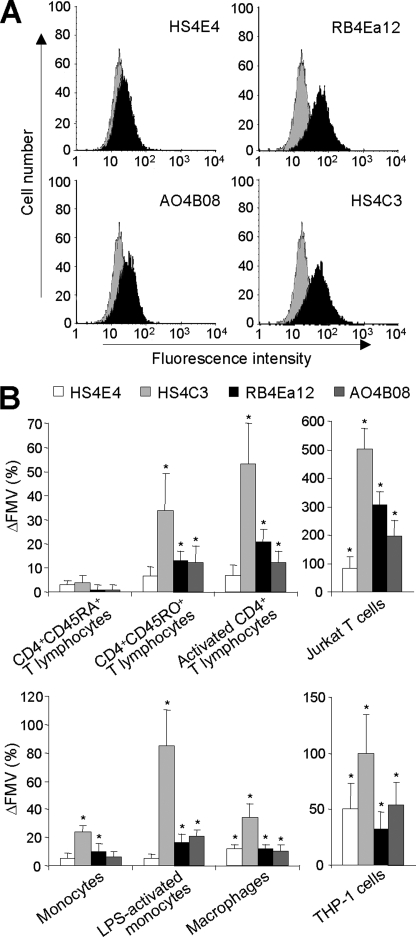
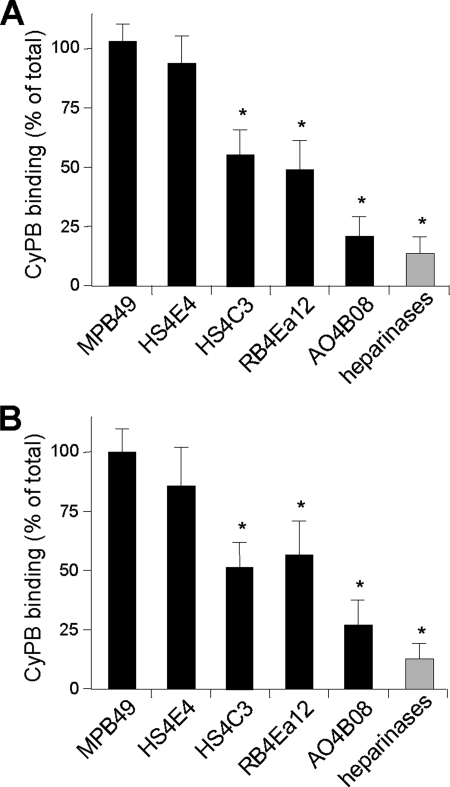
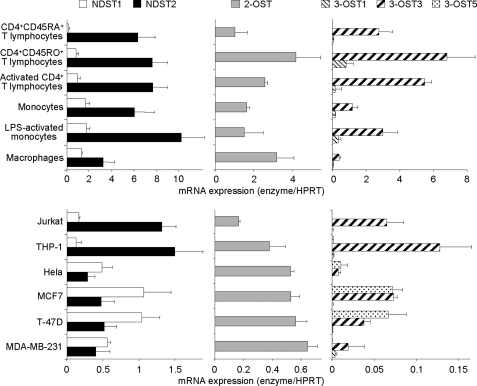
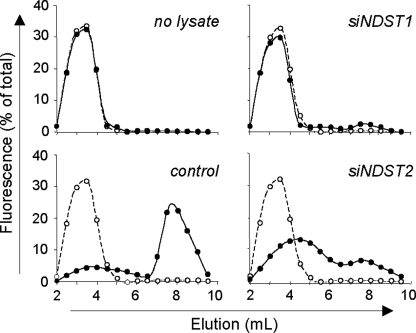
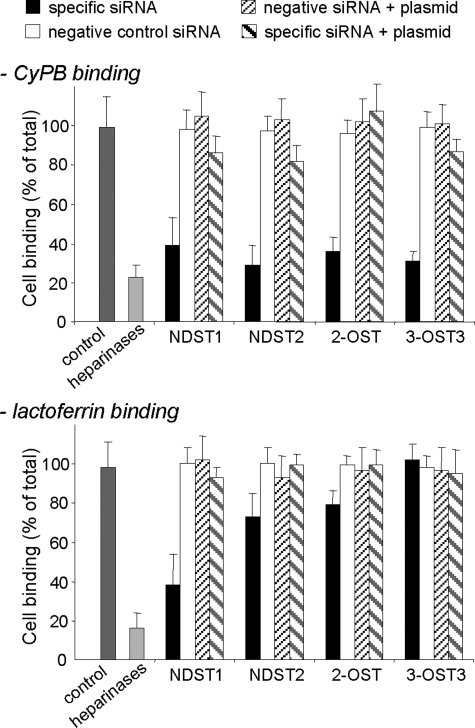
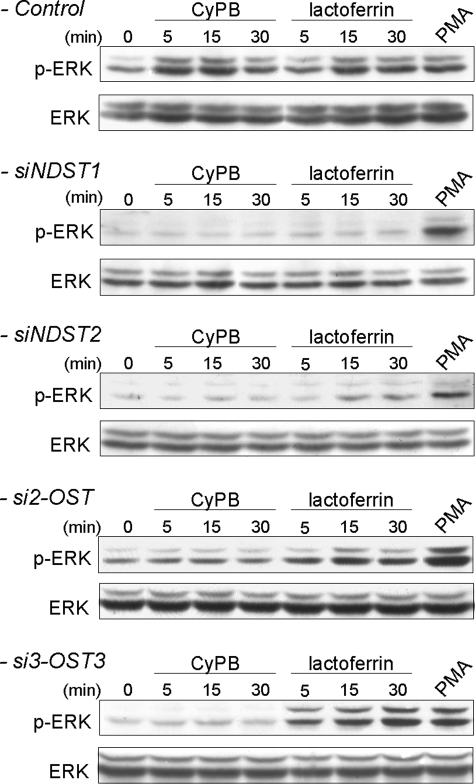
Cyclophilin B (CyPB) induces migration and adhesion of T lymphocytes via a mechanism that requires interaction with 3-O-sulfated heparan sulfate (HS). HS biosynthesis is a complex process with many sulfotransferases involved. N-Deacetylases/N-sulfotransferases are responsible for N-sulfation, which is essential for subsequent modification steps, whereas 3-O-sulfotransferases (3-OSTs) catalyze the least abundant modification. These enzymes are represented by several isoforms, which differ in term of distribution pattern, suggesting their involvement in making tissue-specific HS. To elucidate how the specificity of CyPB binding is determined, we explored the relationships between the expression of these sulfotransferases and the generation of HS motifs with CyPB-binding properties. We demonstrated that high N-sulfate density and the presence of 2-O- and 3-O-sulfates determine binding of CyPB, as evidenced by competitive experiments with heparin derivatives, soluble HS, and anti-HS antibodies. We then showed that target cells, i.e. CD4+ lymphocyte subsets, monocytes/macrophages, and related cell lines, specifically expressed high levels of NDST2 and 3-OST3 isoforms. Silencing the expression of NDST1, NDST2, 2-OST, and 3-OST3 by RNA interference efficiently decreased binding and activity of CyPB, thus confirming their involvement in the biosynthesis of binding sequences for CyPB. Moreover, we demonstrated that NDST1 was able to partially sulfate exogenous substrate in the absence of NDST2 but not vice versa, suggesting that both isoenzymes do not have redundant activities but do have rather complementary activities in making N-sulfated sequences with CyPB-binding properties. Altogether, these results suggest a regulatory mechanism in which cell type-specific expression of certain HS sulfotransferases determines the specific binding of CyPB to target cells.
Figures
Similar articles
-
Heparan sulfate 3-O-sulfation: a rare modification in search of a function.Matrix Biol. 2014 Apr;35:60-72. doi: 10.1016/j.matbio.2013.12.001. Epub 2013 Dec 19. Matrix Biol. 2014. PMID: 24361527 Free PMC article. Review.
-
The heparin/heparan sulfate sequence that interacts with cyclophilin B contains a 3-O-sulfated N-unsubstituted glucosamine residue.J Biol Chem. 2007 Aug 17;282(33):24416-29. doi: 10.1074/jbc.M701835200. Epub 2007 Jun 22. J Biol Chem. 2007. PMID: 17588944
-
Overexpression of heparan sulfate 6-O-sulfotransferases in human embryonic kidney 293 cells results in increased N-acetylglucosaminyl 6-O-sulfation.J Biol Chem. 2006 Mar 3;281(9):5348-56. doi: 10.1074/jbc.M509584200. Epub 2005 Dec 2. J Biol Chem. 2006. PMID: 16326709
-
Enzymatically active N-deacetylase/N-sulfotransferase-2 is present in liver but does not contribute to heparan sulfate N-sulfation.J Biol Chem. 2006 Nov 24;281(47):35727-34. doi: 10.1074/jbc.M604113200. Epub 2006 Sep 19. J Biol Chem. 2006. PMID: 16984905
-
Heparan sulfate and development: differential roles of the N-acetylglucosamine N-deacetylase/N-sulfotransferase isozymes.Biochim Biophys Acta. 2002 Dec 19;1573(3):209-15. doi: 10.1016/s0304-4165(02)00386-0. Biochim Biophys Acta. 2002. PMID: 12417402 Review.
Cited by
-
3-O-Sulfation induces sequence-specific compact topologies in heparan sulfate that encode a dynamic sulfation code.Comput Struct Biotechnol J. 2022 Jul 18;20:3884-3898. doi: 10.1016/j.csbj.2022.07.013. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 35891779 Free PMC article.
-
The heparan sulfate 3-O-sulfotransferases (HS3ST) 2, 3B and 4 enhance proliferation and survival in breast cancer MDA-MB-231 cells.PLoS One. 2018 Mar 16;13(3):e0194676. doi: 10.1371/journal.pone.0194676. eCollection 2018. PLoS One. 2018. PMID: 29547633 Free PMC article.
-
Heparan sulfate 3-O-sulfation: a rare modification in search of a function.Matrix Biol. 2014 Apr;35:60-72. doi: 10.1016/j.matbio.2013.12.001. Epub 2013 Dec 19. Matrix Biol. 2014. PMID: 24361527 Free PMC article. Review.
-
Participation of 3-O-sulfated heparan sulfates in the protection of macrophages by herpes simplex virus-1 glycoprotein D and cyclophilin B against apoptosis.FEBS Open Bio. 2016 Dec 24;7(2):133-148. doi: 10.1002/2211-5463.12145. eCollection 2017 Feb. FEBS Open Bio. 2016. PMID: 28174681 Free PMC article.
-
Sulfotransferase and Heparanase: Remodeling Engines in Promoting Virus Infection and Disease Development.Front Pharmacol. 2018 Nov 22;9:1315. doi: 10.3389/fphar.2018.01315. eCollection 2018. Front Pharmacol. 2018. PMID: 30555321 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
