

Molecular priming of Lyn by GPVI enables an immune receptor to adopt a hemostatic role
- PMID: 19940238
- PMCID: PMC2795544
- DOI: 10.1073/pnas.0906436106
Molecular priming of Lyn by GPVI enables an immune receptor to adopt a hemostatic role
Abstract
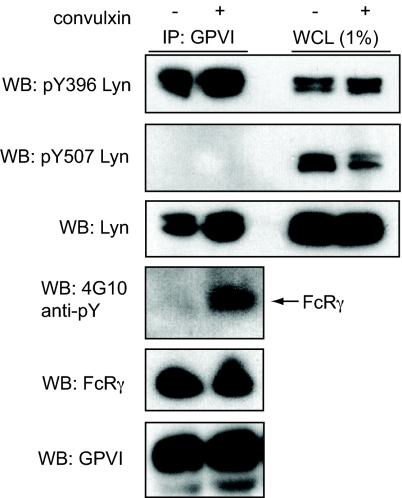
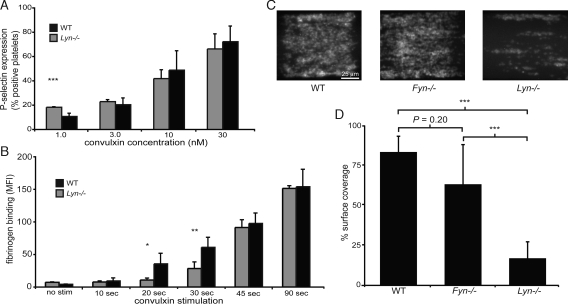
The immune receptor signaling pathway is used by nonimmune cells, but the molecular adaptations that underlie its functional diversification are not known. Circulating platelets use the immune receptor homologue glycoprotein VI (GPVI) to respond to collagen exposed at sites of vessel injury. In contrast to immune cell responses, platelet activation must take place within seconds to successfully form thrombi in flowing blood. Here, we show that the GPVI receptor utilizes a unique intracellular proline-rich domain (PRD) to accelerate platelet activation, a requirement for efficient platelet adhesion to collagen under flow. The GPVI PRD specifically binds the Src-family kinase Lyn and directly activates it, presumably through SH3 displacement. In resting platelets, Lyn is constitutively bound to GPVI in an activated state and platelets lacking Lyn exhibit defective collagen adhesion like that of platelets with GPVI receptors lacking the PRD. These findings define a molecular priming mechanism that enables an immune-type receptor to adopt a hemostatic function. These studies also demonstrate that active kinases can constitutively associate with immune-type receptors without initiating signal transduction before receptor ligation, consistent with a recent molecular model of immune receptor signaling in which receptor ligation is required to bring active kinases to their receptor substrates.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Savage B, Almus-Jacobs F, Ruggeri ZM. Specific synergy of multiple substrate-receptor interactions in platelet thrombus formation under flow. Cell. 1998;94:657–666. - PubMed
-
- Stoll G, Kleinschnitz C, Nieswandt B. Molecular mechanisms of thrombus formation in ischemic stroke: novel insights and targets for treatment. Blood. 2008;112:3555–3562. - PubMed
-
- Clemetson JM, Polgar J, Magnenat E, Wells TN, Clemetson KJ. The platelet collagen receptor glycoprotein VI is a member of the immunoglobulin superfamily closely related to FcalphaR and the natural killer receptors. J Biol Chem. 1999;274:29019–29024. - PubMed
-
- Watson SP, Auger JM, McCarty OJ, Pearce AC. GPVI and integrin alphaIIb beta3 signaling in platelets. J Thromb Haemost. 2005;3:1752–1762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
