

Insights into GATA-1-mediated gene activation versus repression via genome-wide chromatin occupancy analysis
- PMID: 19941827
- PMCID: PMC2800995
- DOI: 10.1016/j.molcel.2009.11.002
Insights into GATA-1-mediated gene activation versus repression via genome-wide chromatin occupancy analysis
Abstract
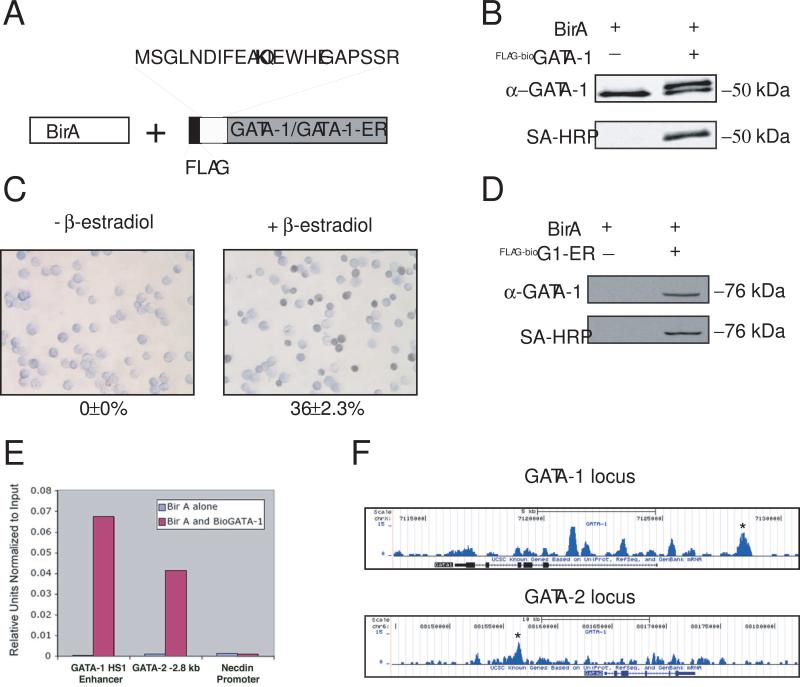
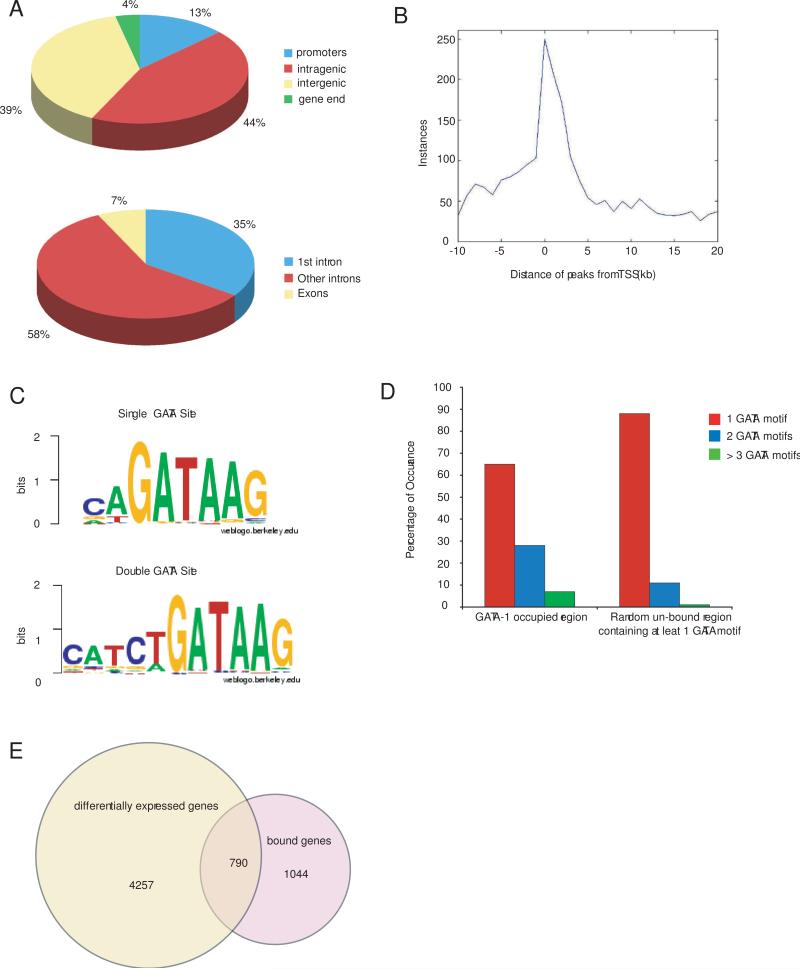
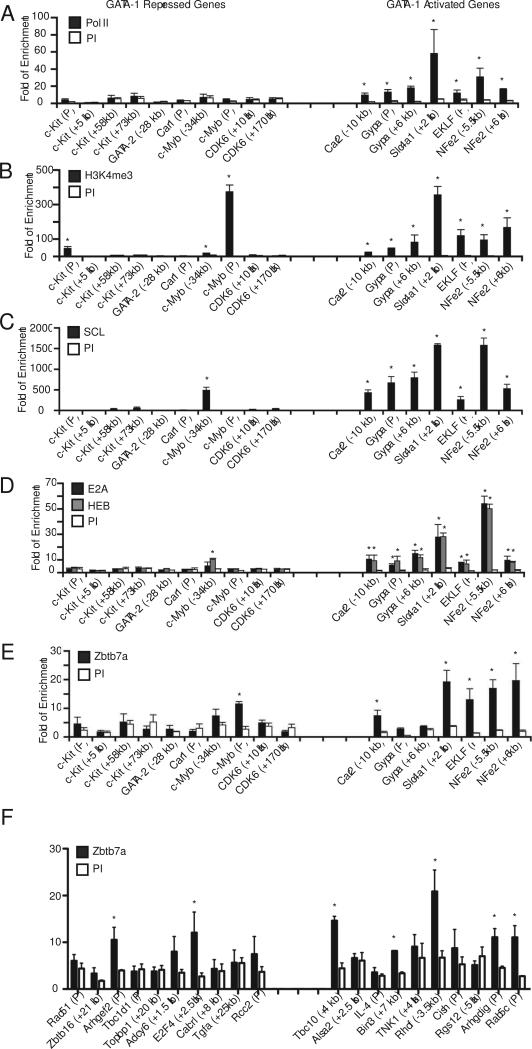
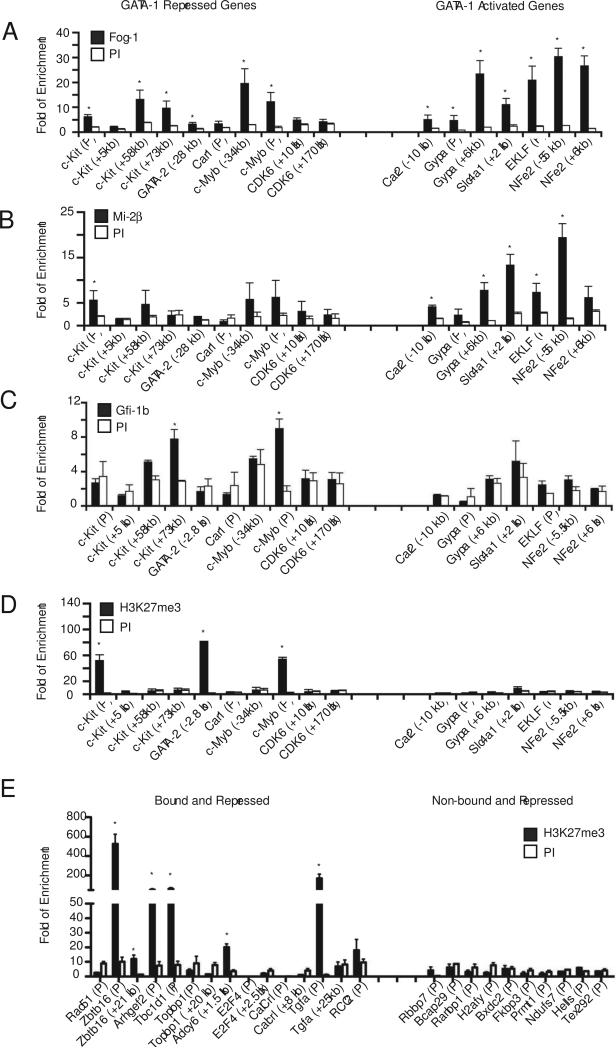
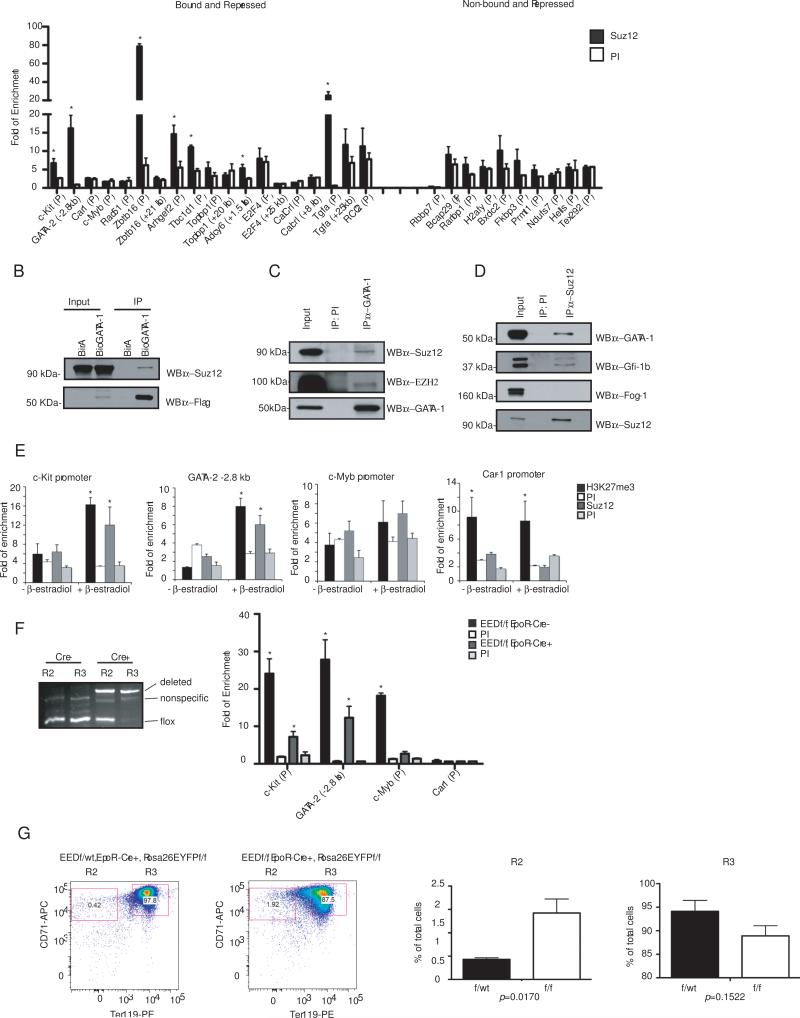
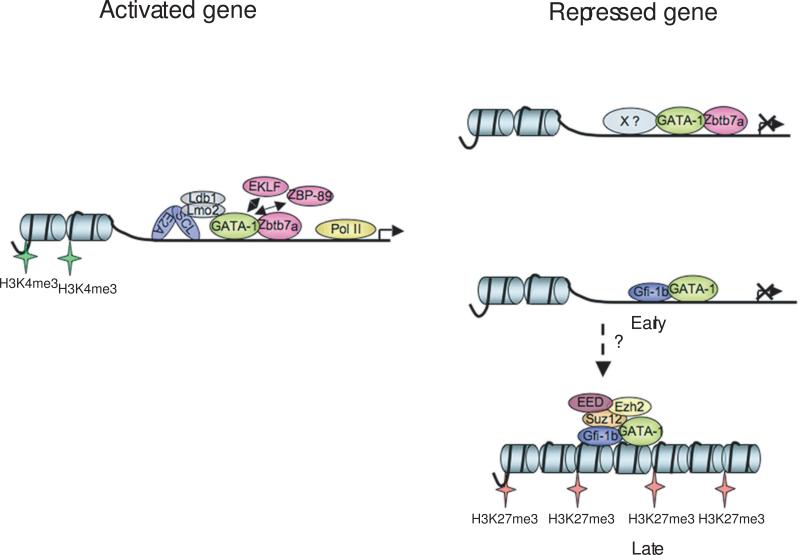
The transcription factor GATA-1 is required for terminal erythroid maturation and functions as an activator or repressor depending on gene context. Yet its in vivo site selectivity and ability to distinguish between activated versus repressed genes remain incompletely understood. In this study, we performed GATA-1 ChIP-seq in erythroid cells and compared it to GATA-1-induced gene expression changes. Bound and differentially expressed genes contain a greater number of GATA-binding motifs, a higher frequency of palindromic GATA sites, and closer occupancy to the transcriptional start site versus nondifferentially expressed genes. Moreover, we show that the transcription factor Zbtb7a occupies GATA-1-bound regions of some direct GATA-1 target genes, that the presence of SCL/TAL1 helps distinguish transcriptional activation versus repression, and that polycomb repressive complex 2 (PRC2) is involved in epigenetic silencing of a subset of GATA-1-repressed genes. These data provide insights into GATA-1-mediated gene regulation in vivo.
Figures
References
-
- Andrews NC, Erdjument-Bromage H, Davidson MB, Tempst P, Orkin SH. Erythroid transcription factor NF-E2 is a haematopoietic-specific basic-leucine zipper protein. Nature. 1993;362:722–728. - PubMed
-
- Bresnick EH, Martowicz ML, Pal S, Johnson KD. Developmental control via GATA factor interplay at chromatin domains. J Cell Physiol. 2005;205:1–9. - PubMed
-
- Cantor AB, Orkin SH. Hematopoietic development: a balancing act. Curr Opin Genet Dev. 2001;11:513–519. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
