

Functional specializations of primary auditory afferents on the Mauthner cells: interactions between membrane and synaptic properties
- PMID: 19941953
- PMCID: PMC3086509
- DOI: 10.1016/j.jphysparis.2009.11.017
Functional specializations of primary auditory afferents on the Mauthner cells: interactions between membrane and synaptic properties
Abstract
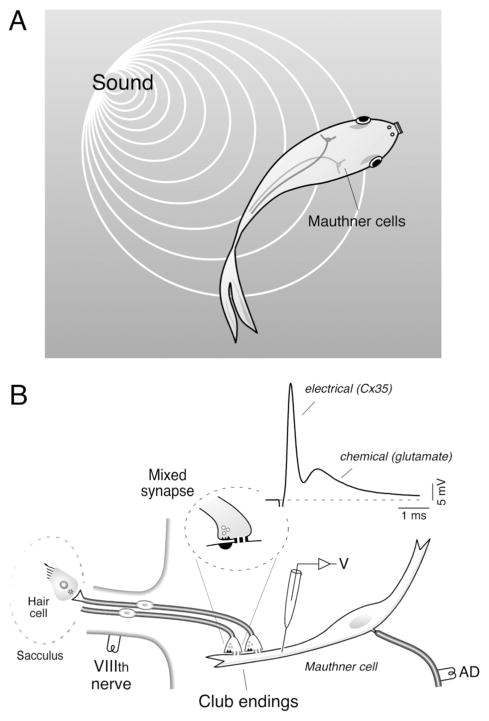
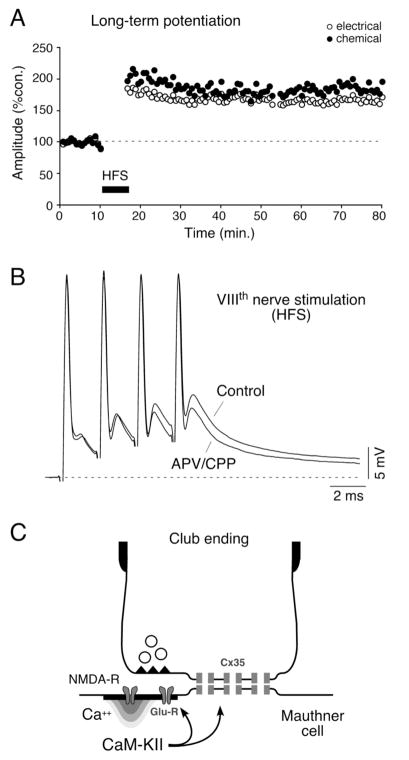
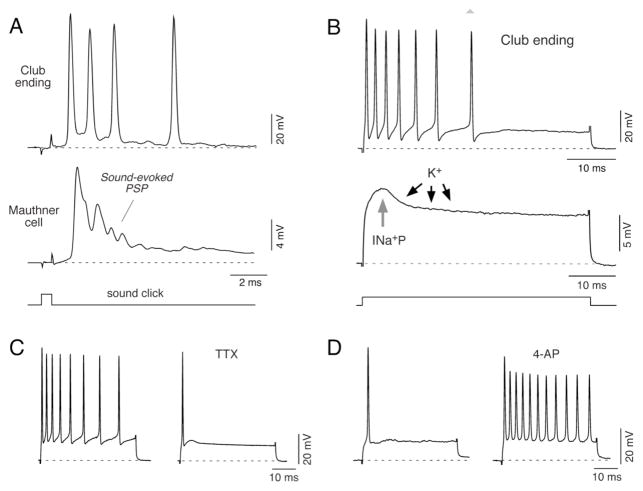
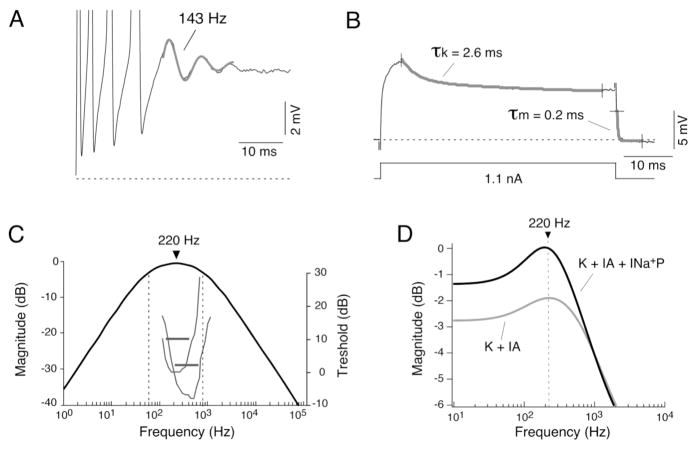
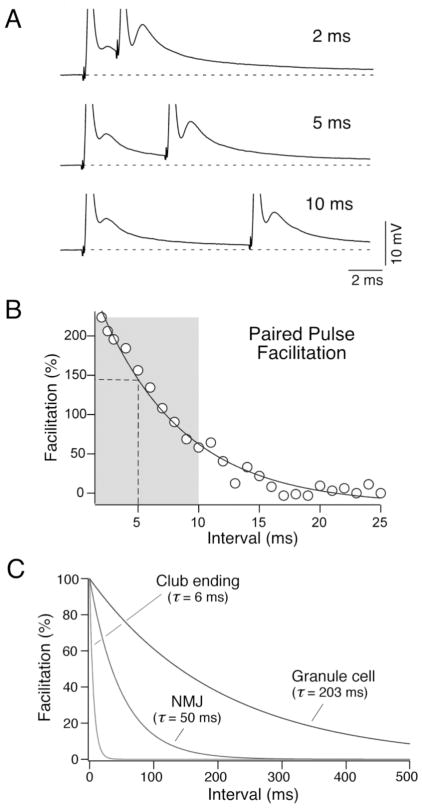
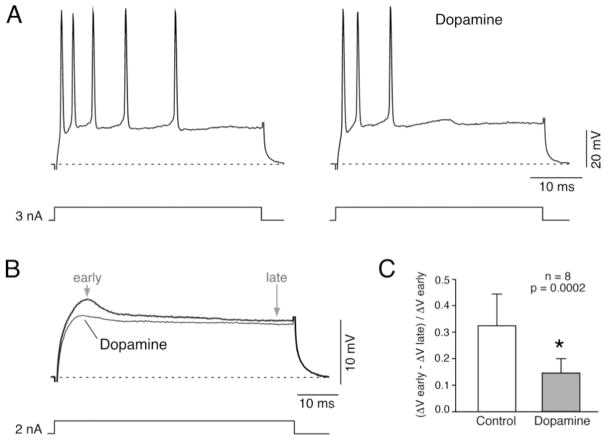
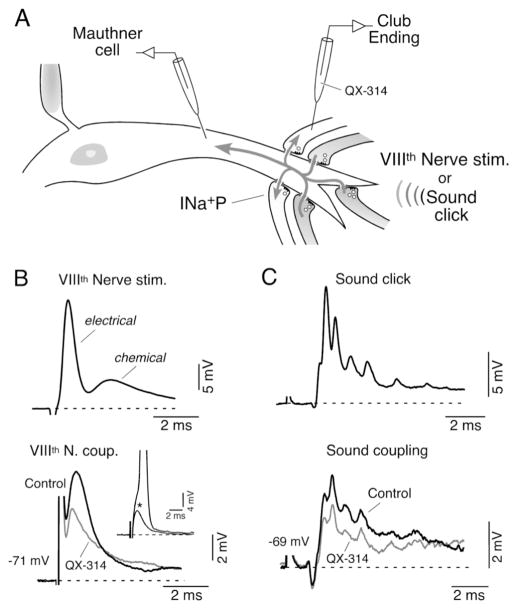
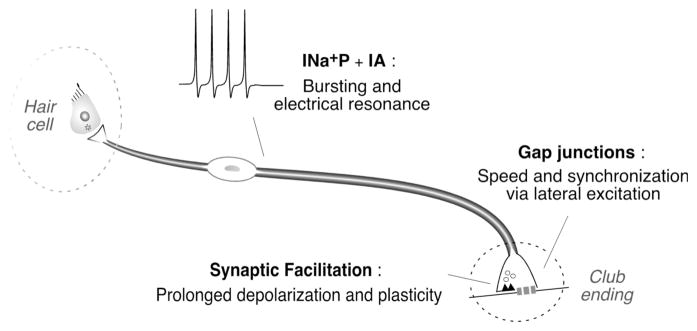
Primary auditory afferents are usually perceived as passive, timing-preserving, lines of communication. Contrasting this view, a special class of auditory afferents to teleost Mauthner cells, a command neuron that organizes tail-flip escape responses, undergoes potentiation of their mixed (electrical and chemical) synapses in response to high frequency cellular activity. This property is likely to represent a mechanism of input sensitization as these neurons provide the Mauthner cell with essential information for the initiation of an escape response. We review here the anatomical and physiological specializations of these identifiable auditory afferents. In particular, we discuss how their membrane and synaptic properties act in concert to more efficaciously activate the Mauthner cells. The striking functional specializations of these neurons suggest that primary auditory afferents might be capable of more sophisticated contributions to auditory processing than has been generally recognized.
Copyright (c) 2009 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Subthreshold sodium current underlies essential functional specializations at primary auditory afferents.J Neurophysiol. 2008 Apr;99(4):1683-99. doi: 10.1152/jn.01173.2007. Epub 2008 Jan 30. J Neurophysiol. 2008. PMID: 18234982
-
Retrograde synaptic communication via gap junctions coupling auditory afferents to the Mauthner cell.J Neurosci. 1995 Sep;15(9):5943-55. doi: 10.1523/JNEUROSCI.15-09-05943.1995. J Neurosci. 1995. PMID: 7666179 Free PMC article.
-
Voltage-dependent enhancement of electrical coupling by a subthreshold sodium current.J Neurosci. 2004 Apr 21;24(16):3999-4010. doi: 10.1523/JNEUROSCI.0077-04.2004. J Neurosci. 2004. PMID: 15102915 Free PMC article.
-
The ion channels and synapses responsible for the physiological diversity of mammalian lower brainstem auditory neurons.Hear Res. 2019 May;376:33-46. doi: 10.1016/j.heares.2018.12.011. Epub 2018 Dec 26. Hear Res. 2019. PMID: 30606624 Review.
-
Synaptic mechanisms for coding timing in auditory neurons.Annu Rev Physiol. 1999;61:477-96. doi: 10.1146/annurev.physiol.61.1.477. Annu Rev Physiol. 1999. PMID: 10099698 Review.
Cited by
-
Electrical synaptic transmission in developing zebrafish: properties and molecular composition of gap junctions at a central auditory synapse.J Neurophysiol. 2014 Nov 1;112(9):2102-13. doi: 10.1152/jn.00397.2014. Epub 2014 Jul 30. J Neurophysiol. 2014. PMID: 25080573 Free PMC article.
-
Cellular Mechanisms of Cortisol-Induced Changes in Mauthner-Cell Excitability in the Startle Circuit of Goldfish.Front Neural Circuits. 2017 Sep 28;11:68. doi: 10.3389/fncir.2017.00068. eCollection 2017. Front Neural Circuits. 2017. PMID: 29033795 Free PMC article.
-
A convergent and essential interneuron pathway for Mauthner-cell-mediated escapes.Curr Biol. 2015 Jun 1;25(11):1526-34. doi: 10.1016/j.cub.2015.04.025. Epub 2015 May 7. Curr Biol. 2015. PMID: 25959971 Free PMC article.
-
Differential processing in modality-specific Mauthner cell dendrites.J Physiol. 2018 Feb 15;596(4):667-689. doi: 10.1113/JP274861. Epub 2017 Dec 18. J Physiol. 2018. PMID: 29148564 Free PMC article.
-
Two independent forms of activity-dependent potentiation regulate electrical transmission at mixed synapses on the Mauthner cell.Brain Res. 2012 Dec 3;1487:173-82. doi: 10.1016/j.brainres.2012.05.059. Epub 2012 Jul 4. Brain Res. 2012. PMID: 22771708 Free PMC article. Review.
References
-
- Bartelmez GW. Mauthner’s cell and the nucleus motorius tegmenti. J Comp Neurol. 1915;25:87–128.
-
- Beccari N. Richerche sulle cellule e fibre del Mauthner e sulle loro conessioni in pesci ed anfibii. Arch Ital Anay E Embr. 1907;T.6:660–705.
-
- Bodian D. The structure of the vertebrate synapse. A study of the axon endings on Mauthner’s cell and neighboring centers in the goldfish. J Comp Neurol. 1937;1:117–160.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
