

Mechanism of action of bolandiol (19-nortestosterone-3beta,17beta-diol), a unique anabolic steroid with androgenic, estrogenic, and progestational activities
- PMID: 19941958
- PMCID: PMC2831543
- DOI: 10.1016/j.jsbmb.2009.11.008
Mechanism of action of bolandiol (19-nortestosterone-3beta,17beta-diol), a unique anabolic steroid with androgenic, estrogenic, and progestational activities
Abstract
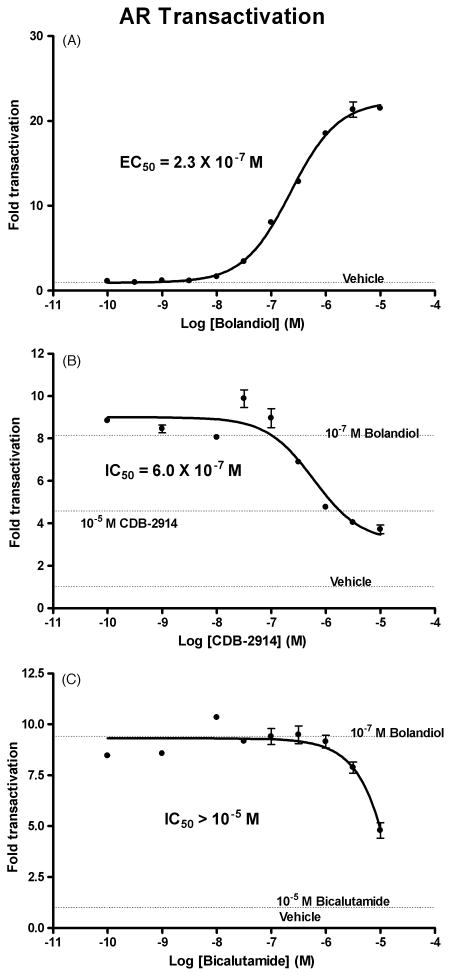
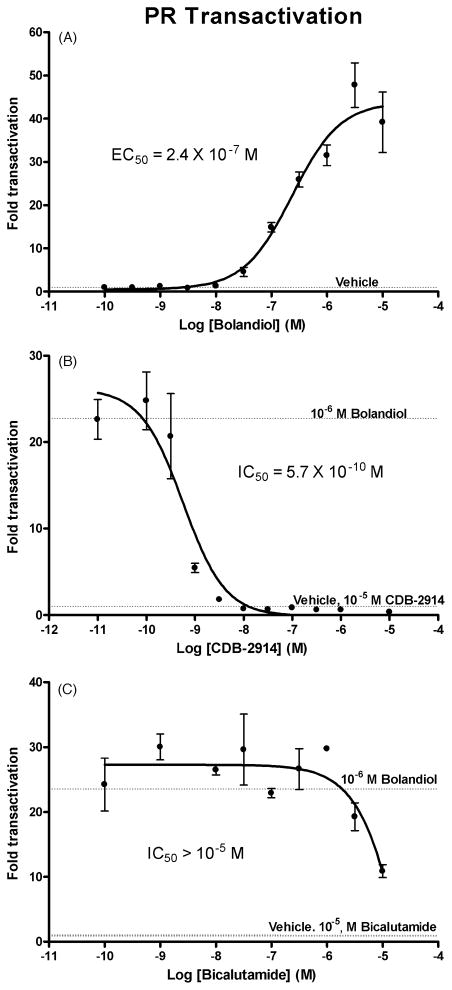
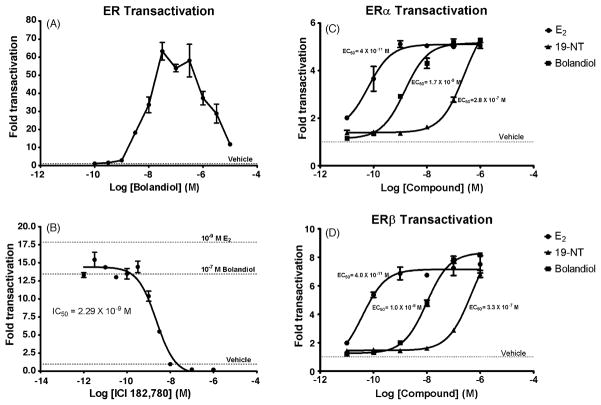
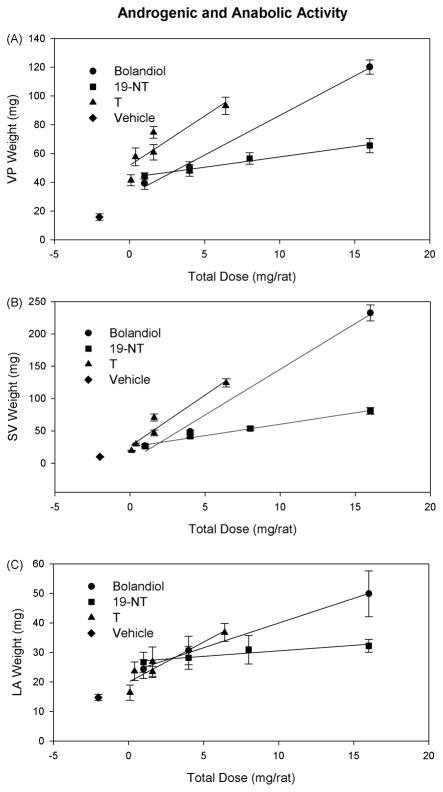
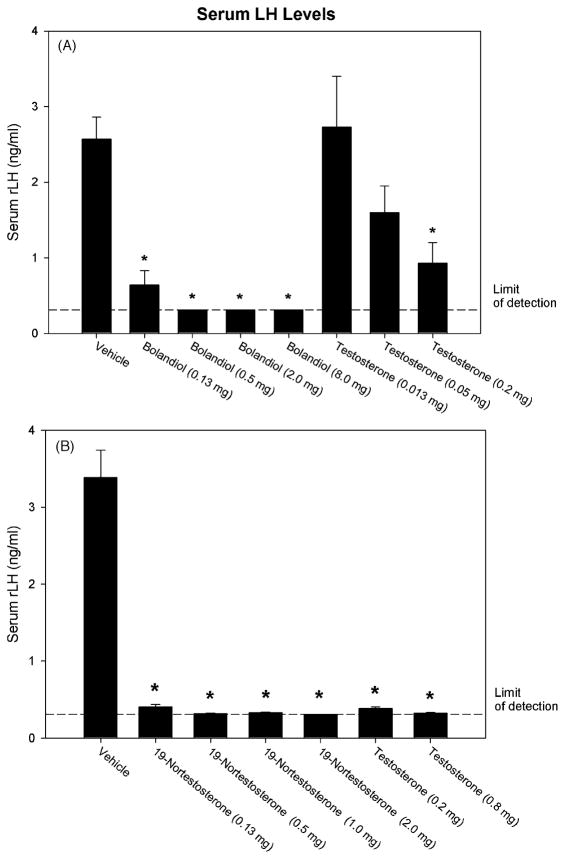
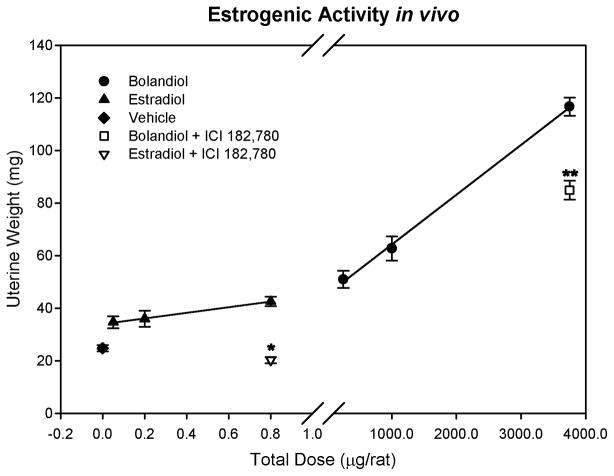
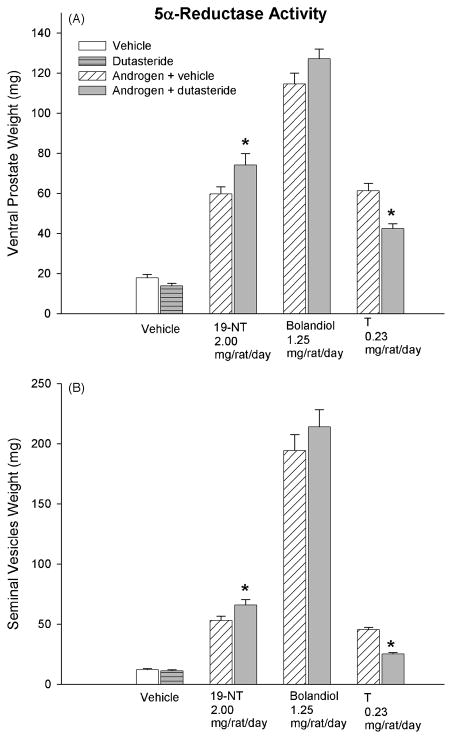
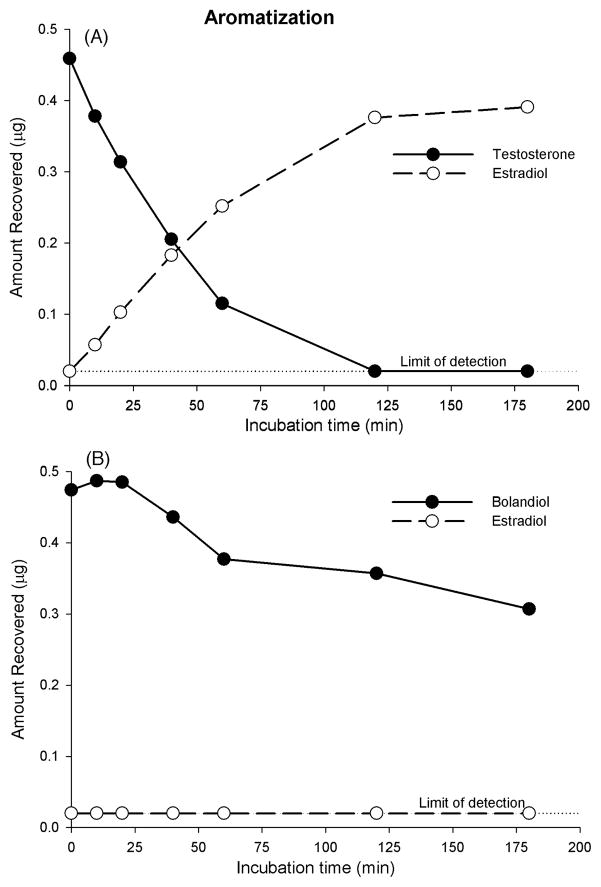
Bolandiol is a synthetic anabolic steroid that increases lean body mass and bone mineral density without significant stimulation of sex accessory glands in castrate adult male rats. Since bolandiol suppresses gonadotropins and endogenous testosterone (T) production, we investigated its mechanism of action. We compared the potency of bolandiol in vitro and in vivo with T, 5alpha-dihydrotestosterone (DHT), 19-nortestosterone (19-NT) and estradiol (E(2)). Bolandiol bound with lower affinity to the recombinant rat androgen receptor (AR) than the other androgens and had low, but measurable, affinity for recombinant human progestin receptors (PR-A, PR-B), and estrogen receptors (ERalpha and beta-1). Functional agonist activity was assessed in transcription assays mediated by AR, PR, or ER. Bolandiol was stimulatory in all these assays, but only 4-9% as potent as T, DHT, and 19-NT via AR, 1% as potent as progesterone via PR, and 3% and 1% as potent as E(2) acting through ERalpha or ERbeta, respectively. In immature castrate rats, bolandiol was equipotent to T in stimulating growth of the levator ani muscle but less potent than T in stimulating growth of the sex accessory glands. Bolandiol also stimulated uterine weight increases in immature female rats, which were partly blocked by ICI 182,780, but it was not aromatized in vitro by recombinant human aromatase. In contrast to T, stimulation of sex accessory gland weights by bolandiol was not inhibited by concomitant treatment with the dual 5alpha-reductase inhibitor dutasteride. As bolandiol exhibits tissue selectivity in vivo, it may act via AR, PR, and/or ER, utilize alternative signaling pathway(s) or transcriptional coregulators, and/or be metabolized to a more potent selective steroid.
Figures
Similar articles
-
The potent synthetic androgens, dimethandrolone (7α,11β-dimethyl-19-nortestosterone) and 11β-methyl-19-nortestosterone, do not require 5α-reduction to exert their maximal androgenic effects.J Steroid Biochem Mol Biol. 2010 Oct;122(4):212-8. doi: 10.1016/j.jsbmb.2010.06.009. Epub 2010 Jun 25. J Steroid Biochem Mol Biol. 2010. PMID: 20599615 Free PMC article.
-
Estrogenic and progestational activity of 7alpha-methyl-19-nortestosterone, a synthetic androgen.J Steroid Biochem Mol Biol. 1998 Nov;67(3):275-83. doi: 10.1016/s0960-0760(98)00114-9. J Steroid Biochem Mol Biol. 1998. PMID: 9879986
-
Long-term effects of dimethandrolone 17β-undecanoate and 11β-methyl-19-nortestosterone 17β-dodecylcarbonate on body composition, bone mineral density, serum gonadotropins, and androgenic/anabolic activity in castrated male rats.J Androl. 2011 Mar-Apr;32(2):183-92. doi: 10.2164/jandrol.110.010371. Epub 2010 Aug 26. J Androl. 2011. PMID: 20798389
-
Preclinical evaluation of norgestimate, a progestin with minimal androgenic activity.Am J Obstet Gynecol. 1992 Oct;167(4 Pt 2):1191-6. doi: 10.1016/s0002-9378(12)90410-x. Am J Obstet Gynecol. 1992. PMID: 1415445 Review.
-
Prolactin influences upon androgen action in male accessory sex organs.Adv Sex Horm Res. 1976;2:425-70. Adv Sex Horm Res. 1976. PMID: 189591 Review.
Cited by
-
Enhanced evaluation of selective androgen receptor modulators in vivo.Andrology. 2013 Jan;1(1):29-36. doi: 10.1111/j.2047-2927.2012.00006.x. Epub 2012 Aug 28. Andrology. 2013. PMID: 23258627 Free PMC article.
-
Modulatory effect of gonadotropins on rats' ovaries after nandrolone decanoate administration: a stereological study.Iran J Med Sci. 2014 Jan;39(1):44-50. Iran J Med Sci. 2014. PMID: 24453393 Free PMC article.
-
The Buzz about anabolic androgenic steroids: electrophysiological effects in excitable tissues.Neuroendocrinology. 2012;96(2):141-51. doi: 10.1159/000339123. Epub 2012 Sep 14. Neuroendocrinology. 2012. PMID: 22576754 Free PMC article. Review.
-
In vitro androgen bioassays as a detection method for designer androgens.Sensors (Basel). 2013 Feb 6;13(2):2148-63. doi: 10.3390/s130202148. Sensors (Basel). 2013. PMID: 23389345 Free PMC article. Review.
-
Anabolic androgenic steroid abuse: multiple mechanisms of regulation of GABAergic synapses in neuroendocrine control regions of the rodent forebrain.J Neuroendocrinol. 2012 Jan;24(1):202-14. doi: 10.1111/j.1365-2826.2011.02151.x. J Neuroendocrinol. 2012. PMID: 21554430 Free PMC article. Review.
References
-
- Schrader Y, Thevis M, Schanzer W. Quantitative determination of metabolic products of 19-norandrostenediol in human plasma using gas chromatography/mass spectrometry. Drug Metab Dispos. 2006;34:1328–1335. - PubMed
-
- Page ST, Marck BT, Tolliver JM, Matsumoto AM. Tissue selectivity of the anabolic steroid, 19-nor-4-androstenediol-3beta, 17beta-diol in male Sprague Dawley rats: selective stimulation of muscle mass and bone mineral density relative to prostate mass. Endocrinology. 2008;149:1987–1993. - PMC - PubMed
-
- Van Gammeren D, Falk D, Antonio J. The effects of supplementation with 19-nor-4-androstene-3, 17-dione and 19-nor-4-androstene-3, 17-diol on body composition and athletic performance in previously weight-trained male athletes. Eur J Appl Physiol. 2001;84:426–431. - PubMed
-
- Van Gammeren D, Falk D, Antonio J. Effects of norandrostenedione and norandrostenediol in resistance-trained men. Nutrition. 2002;18:734–737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
