

Molecular architecture of Streptococcus pneumoniae TIGR4 pili
- PMID: 19942854
- PMCID: PMC2797065
- DOI: 10.1038/emboj.2009.360
Molecular architecture of Streptococcus pneumoniae TIGR4 pili
Abstract
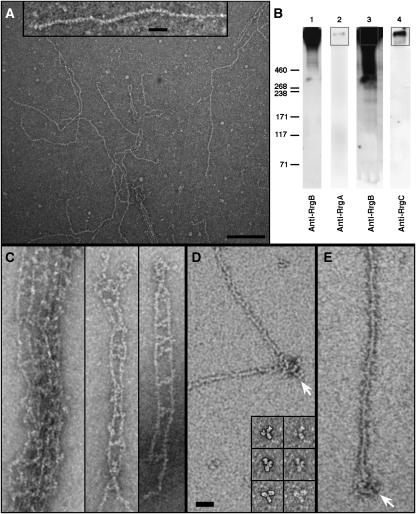
Although the pili of Gram-positive bacteria are putative virulence factors, little is known about their structure. Here we describe the molecular architecture of pilus-1 of Streptococcus pneumoniae, which is a major cause of morbidity and mortality worldwide. One major (RrgB) and two minor components (RrgA and RrgC) assemble into the pilus. Results from TEM and scanning transmission EM show that the native pili are approximately 6 nm wide, flexible filaments that can be over 1 microm long. They are formed by a single string of RrgB monomers and have a polarity defined by nose-like protrusions. These protrusions correlate to the shape of monomeric RrgB-His, which like RrgA-His and RrgC-His has an elongated, multi-domain structure. RrgA and RrgC are only present at the opposite ends of the pilus shaft, compatible with their putative roles as adhesin and anchor to the cell wall surface, respectively. Our structural analyses provide the first direct experimental evidence that the native S. pneumoniae pilus shaft is composed exclusively of covalently linked monomeric RrgB subunits oriented head-to-tail.
Conflict of interest statement
MH, GD, RR and IF are employees of Novartis Vaccines and Diagnostics s.r.l.
Figures





Similar articles
-
Association of RrgA and RrgC into the Streptococcus pneumoniae pilus by sortases C-2 and C-3.Biochemistry. 2012 Jan 10;51(1):342-52. doi: 10.1021/bi201591n. Epub 2011 Dec 7. Biochemistry. 2012. PMID: 22122269
-
Roles of the sortases of Streptococcus pneumoniae in assembly of the RlrA pilus.J Bacteriol. 2008 Sep;190(17):6002-13. doi: 10.1128/JB.00379-08. Epub 2008 Jul 7. J Bacteriol. 2008. PMID: 18606733 Free PMC article.
-
Supramolecular organization of the repetitive backbone unit of the Streptococcus pneumoniae pilus.PLoS One. 2010 Jun 15;5(6):e10919. doi: 10.1371/journal.pone.0010919. PLoS One. 2010. PMID: 20559564 Free PMC article.
-
Pilus of Streptococcus pneumoniae: structure, function and vaccine potential.Front Cell Infect Microbiol. 2023 Sep 20;13:1270848. doi: 10.3389/fcimb.2023.1270848. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37799336 Free PMC article. Review.
-
The impact of the ancillary pilus-1 protein RrgA of Streptococcus pneumoniae on colonization and disease.Mol Microbiol. 2020 Mar;113(3):650-658. doi: 10.1111/mmi.14451. Mol Microbiol. 2020. PMID: 32185835 Review.
Cited by
-
The Streptococcus pneumoniae pilus-1 displays a biphasic expression pattern.PLoS One. 2011;6(6):e21269. doi: 10.1371/journal.pone.0021269. Epub 2011 Jun 22. PLoS One. 2011. PMID: 21731688 Free PMC article.
-
Protocol for pneumococcal meningitis induction in mice via non-surgical intracisternal injection.STAR Protoc. 2025 Jun 20;6(2):103773. doi: 10.1016/j.xpro.2025.103773. Epub 2025 Apr 19. STAR Protoc. 2025. PMID: 40253700 Free PMC article.
-
Understanding pneumococcal serotype 1 biology through population genomic analysis.BMC Infect Dis. 2016 Nov 8;16(1):649. doi: 10.1186/s12879-016-1987-z. BMC Infect Dis. 2016. PMID: 27821148 Free PMC article.
-
The ancillary protein 1 of Streptococcus pyogenes FCT-1 pili mediates cell adhesion and biofilm formation through heterophilic as well as homophilic interactions.Mol Microbiol. 2012 Mar;83(5):1035-47. doi: 10.1111/j.1365-2958.2012.07987.x. Epub 2012 Feb 10. Mol Microbiol. 2012. PMID: 22320452 Free PMC article.
-
Unencapsulated Streptococcus pneumoniae from conjunctivitis encode variant traits and belong to a distinct phylogenetic cluster.Nat Commun. 2014 Nov 12;5:5411. doi: 10.1038/ncomms6411. Nat Commun. 2014. PMID: 25388376 Free PMC article.
References
-
- Abbot EL, Smith WD, Siou GP, Chiriboga C, Smith RJ, Wilson JA, Hirst BH, Kehoe MA (2007) Pili mediate specific adhesion of Streptococcus pyogenes to human tonsil and skin. Cell Microbiol 9: 1822–1833 - PubMed
-
- Bagnoli F, Moschioni M, Donati C, Dimitrovska V, Ferlenghi I, Facciotti C, Muzzi A, Giusti F, Emolo C, Sinisi A, Hilleringmann M, Pansegrau W, Censini S, Rappuoli R, Covacci A, Masignani V, Barocchi MA (2008) A second pilus type in Streptococcus pneumoniae is prevalent in emerging serotypes and mediates adhesion to host cells. J Bacteriol 190: 5480–5492 - PMC - PubMed
-
- Barocchi MA, Ries J, Zogaj X, Hemsley C, Albiger B, Kanth A, Dahlberg S, Fernebro J, Moschioni M, Masignani V, Hultenby K, Taddei AR, Beiter K, Wartha F, von Euler A, Covacci A, Holden DW, Normark S, Rappuoli R, Henriques-Normark B (2006) A pneumococcal pilus influences virulence and host inflammatory responses. Proc Natl Acad Sci USA 103: 2857–2862 - PMC - PubMed
-
- Broz P, Mueller CA, Muller SA, Philippsen A, Sorg I, Engel A, Cornelis GR (2007) Function and molecular architecture of the Yersinia injectisome tip complex. Mol Microbiol 65: 1311–1320 - PubMed
-
- Budzik JM, Marraffini LA, Schneewind O (2007) Assembly of pili on the surface of Bacillus cereus vegetative cells. Mol Microbiol 66: 495–510 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
