

Analysis of small RNA in fission yeast; centromeric siRNAs are potentially generated through a structured RNA
- PMID: 19942857
- PMCID: PMC2797062
- DOI: 10.1038/emboj.2009.351
Analysis of small RNA in fission yeast; centromeric siRNAs are potentially generated through a structured RNA
Abstract
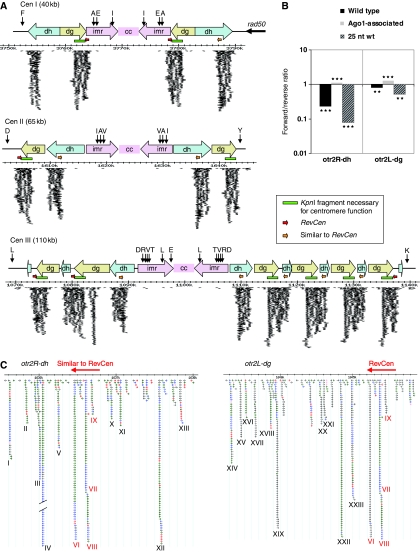
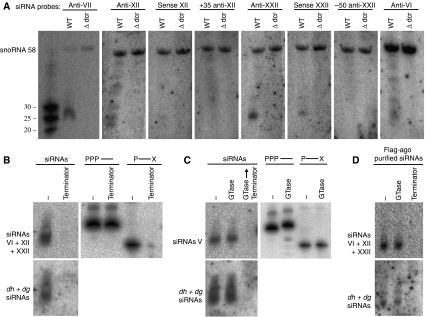
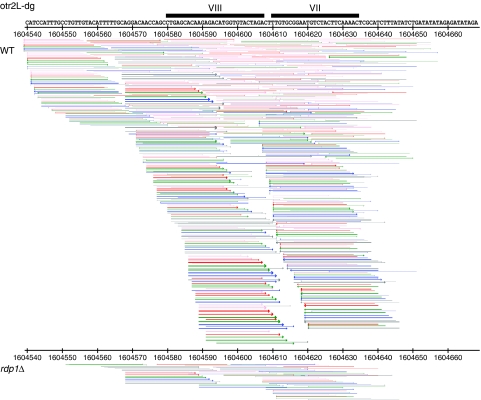
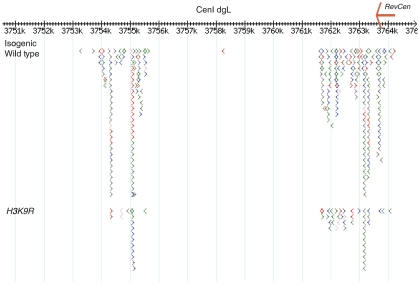
The formation of heterochromatin at the centromeres in fission yeast depends on transcription of the outer repeats. These transcripts are processed into siRNAs that target homologous loci for heterochromatin formation. Here, high throughput sequencing of small RNA provides a comprehensive analysis of centromere-derived small RNAs. We found that the centromeric small RNAs are Dcr1 dependent, carry 5'-monophosphates and are associated with Ago1. The majority of centromeric small RNAs originate from two remarkably well-conserved sequences that are present in all centromeres. The high degree of similarity suggests that this non-coding sequence in itself may be of importance. Consistent with this, secondary structure-probing experiments indicate that this centromeric RNA is partially double-stranded and is processed by Dicer in vitro. We further demonstrate the existence of small centromeric RNA in rdp1Delta cells. Our data suggest a pathway for siRNA generation that is distinct from the well-documented model involving RITS/RDRC. We propose that primary transcripts fold into hairpin-like structures that may be processed by Dcr1 into siRNAs, and that these siRNAs may initiate heterochromatin formation independent of RDRC activity.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Similar articles
-
Coupling of double-stranded RNA synthesis and siRNA generation in fission yeast RNAi.Mol Cell. 2007 Aug 3;27(3):449-61. doi: 10.1016/j.molcel.2007.07.007. Epub 2007 Jul 19. Mol Cell. 2007. PMID: 17658285
-
Chp1-Tas3 interaction is required to recruit RITS to fission yeast centromeres and for maintenance of centromeric heterochromatin.Mol Cell Biol. 2008 Apr;28(7):2154-66. doi: 10.1128/MCB.01637-07. Epub 2008 Jan 22. Mol Cell Biol. 2008. PMID: 18212052 Free PMC article.
-
Studies on the mechanism of RNAi-dependent heterochromatin assembly.Cold Spring Harb Symp Quant Biol. 2006;71:461-71. doi: 10.1101/sqb.2006.71.044. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381328 Review.
-
RNA interference (RNAi)-dependent and RNAi-independent association of the Chp1 chromodomain protein with distinct heterochromatic loci in fission yeast.Mol Cell Biol. 2005 Mar;25(6):2331-46. doi: 10.1128/MCB.25.6.2331-2346.2005. Mol Cell Biol. 2005. PMID: 15743828 Free PMC article.
-
On the connection between RNAi and heterochromatin at centromeres.Cold Spring Harb Symp Quant Biol. 2010;75:275-83. doi: 10.1101/sqb.2010.75.024. Epub 2011 Feb 2. Cold Spring Harb Symp Quant Biol. 2010. PMID: 21289046 Review.
Cited by
-
Tandemly repeated genes promote RNAi-mediated heterochromatin formation via an antisilencing factor, Epe1, in fission yeast.Genes Dev. 2022 Nov-Dec 1;36(21-24):1145-1159. doi: 10.1101/gad.350129.122. Epub 2022 Dec 20. Genes Dev. 2022. PMID: 36617881 Free PMC article.
-
Epigenetic regulation of centromere function.Cell Mol Life Sci. 2020 Aug;77(15):2899-2917. doi: 10.1007/s00018-020-03460-8. Epub 2020 Feb 1. Cell Mol Life Sci. 2020. PMID: 32008088 Free PMC article. Review.
-
Genome-wide identification of allele-specific expression in response to Streptococcus suis 2 infection in two differentially susceptible pig breeds.J Appl Genet. 2015 Nov;56(4):481-491. doi: 10.1007/s13353-015-0275-8. Epub 2015 Mar 4. J Appl Genet. 2015. PMID: 25737137
-
Conserved chromosomal functions of RNA interference.Nat Rev Genet. 2020 May;21(5):311-331. doi: 10.1038/s41576-019-0203-6. Epub 2020 Feb 12. Nat Rev Genet. 2020. PMID: 32051563 Free PMC article. Review.
-
RNAi-dependent formation of heterochromatin and its diverse functions.Curr Opin Genet Dev. 2010 Apr;20(2):134-41. doi: 10.1016/j.gde.2010.02.003. Epub 2010 Mar 6. Curr Opin Genet Dev. 2010. PMID: 20207534 Free PMC article. Review.
References
-
- Allshire RC, Nimmo ER, Ekwall K, Javerzat JP, Cranston G (1995) Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev 9: 218–233 - PubMed
-
- Appelgren H, Kniola B, Ekwall K (2003) Distinct centromere domain structures with separate functions demonstrated in live fission yeast cells. J Cell Sci 116 (Pt 19): 4035–4042 - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T (2001) Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410: 120–124 - PubMed
-
- Bernard P, Maure JF, Partridge JF, Genier S, Javerzat JP, Allshire RC (2001) Requirement of heterochromatin for cohesion at centromeres. Science 294: 2539–2542 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
