

A vestibular sensation: probabilistic approaches to spatial perception
- PMID: 19945388
- PMCID: PMC2786181
- DOI: 10.1016/j.neuron.2009.11.010
A vestibular sensation: probabilistic approaches to spatial perception
Abstract
The vestibular system helps maintain equilibrium and clear vision through reflexes, but it also contributes to spatial perception. In recent years, research in the vestibular field has expanded to higher-level processing involving the cortex. Vestibular contributions to spatial cognition have been difficult to study because the circuits involved are inherently multisensory. Computational methods and the application of Bayes theorem are used to form hypotheses about how information from different sensory modalities is combined together with expectations based on past experience in order to obtain optimal estimates of cognitive variables like current spatial orientation. To test these hypotheses, neuronal populations are being recorded during active tasks in which subjects make decisions based on vestibular and visual or somatosensory information. This review highlights what is currently known about the role of vestibular information in these processes, the computations necessary to obtain the appropriate signals, and the benefits that have emerged thus far.
Figures
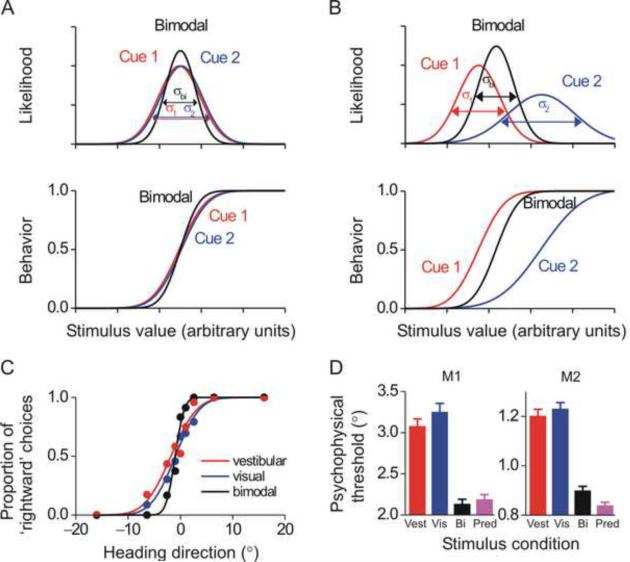
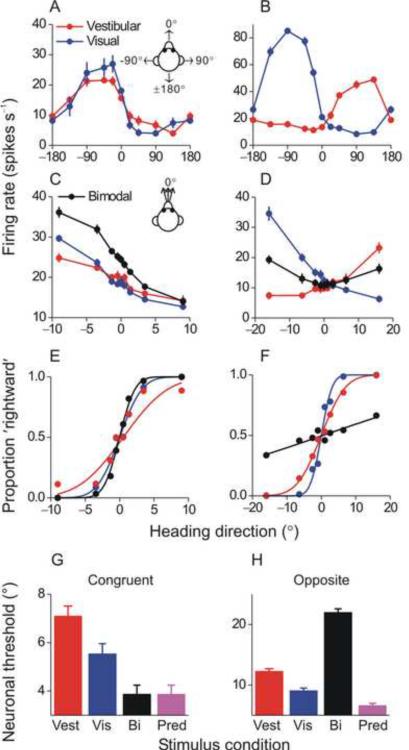
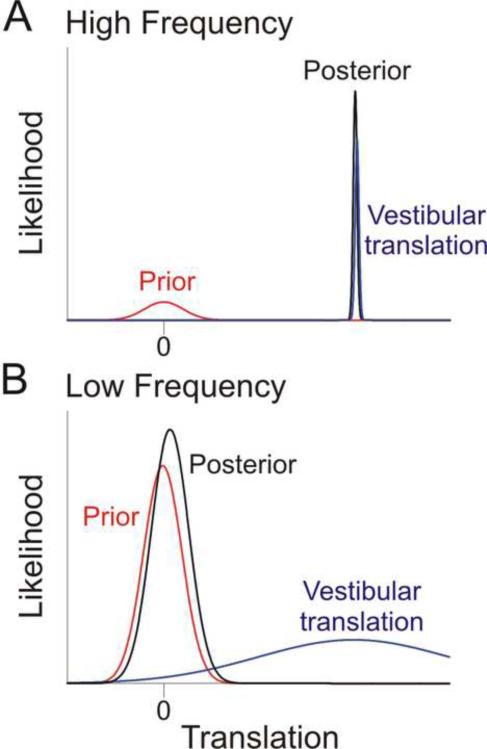
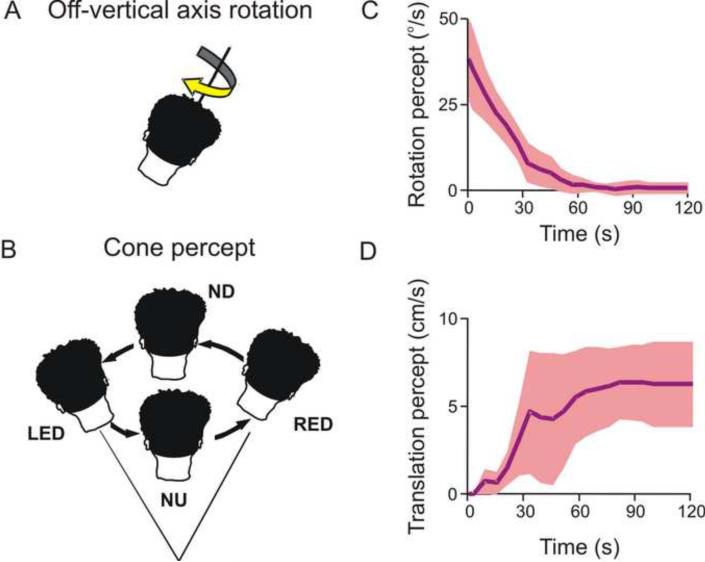
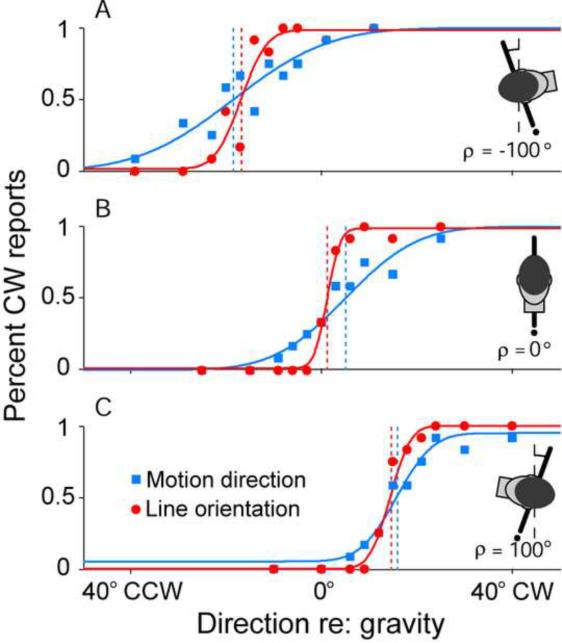
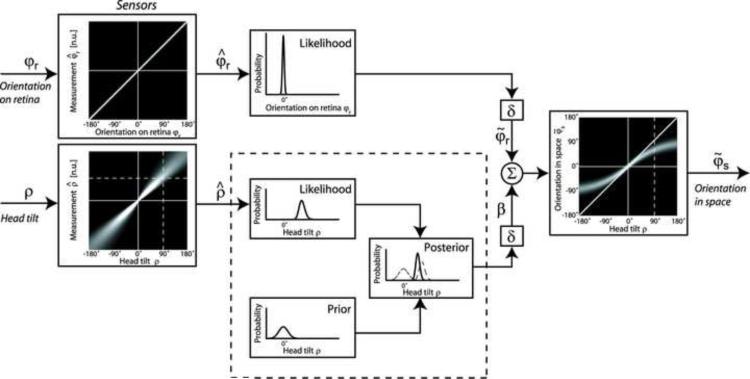
References
-
- Angelaki DE. Three-dimensional organization of otolith-ocular reflexes in rhesus monkeys. III. Responses to translation. J. Neurophysiol. 1998;80:680–695. - PubMed
-
- Angelaki DE. Eyes on target: what neurons must do for the vestibuloocular reflex during linear motion. J. Neurophysiol. 2004;92:20–35. - PubMed
-
- Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodal sense. Annu. Rev. Neurosci. 2008;31:125–150. - PubMed
-
- Angelaki DE, Hess BJ. Inertial representation of angular motion in the vestibular system of rhesus monkeys. II. Otolith-controlled transformation that depends on an intact cerebellar nodulus. J. Neurophysiol. 1995a;73:1729–1751. - PubMed
-
- Angelaki DE, Hess BJ. Lesion of the nodulus and ventral uvula abolish steady-state off-vertical axis otolith response. J. Neurophysiol. 1995b;73:1716–1720. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
