

Classical MHCI molecules regulate retinogeniculate refinement and limit ocular dominance plasticity
- PMID: 19945389
- PMCID: PMC2787480
- DOI: 10.1016/j.neuron.2009.10.015
Classical MHCI molecules regulate retinogeniculate refinement and limit ocular dominance plasticity
Abstract
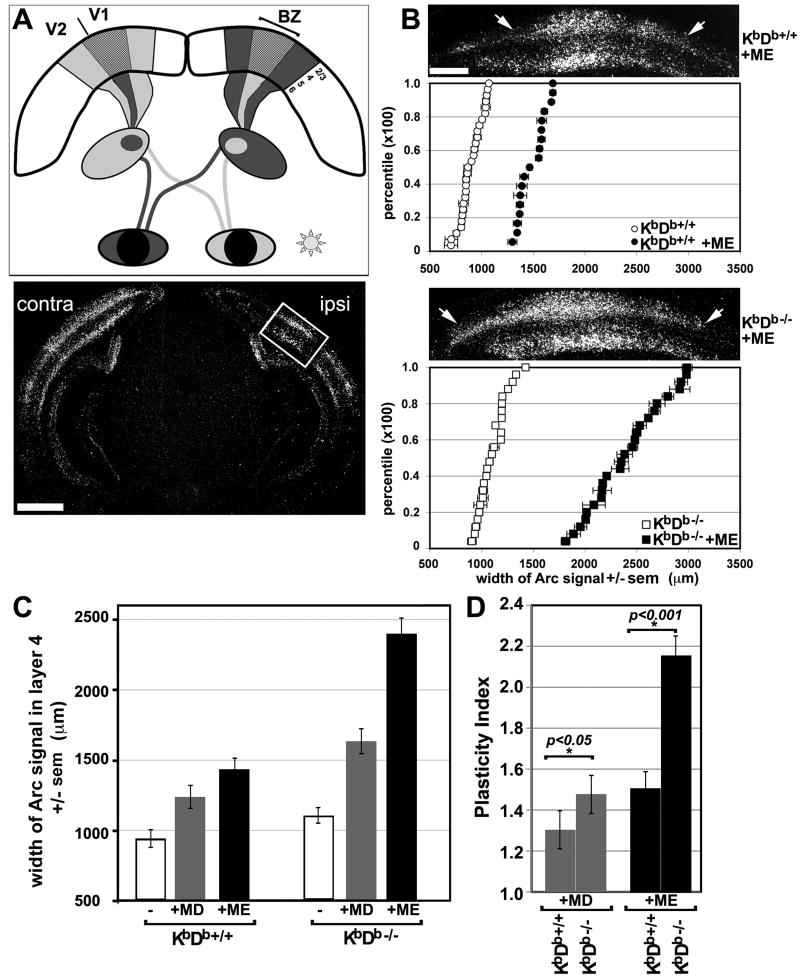
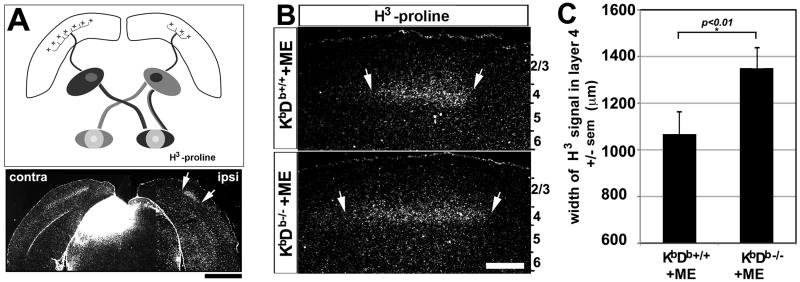
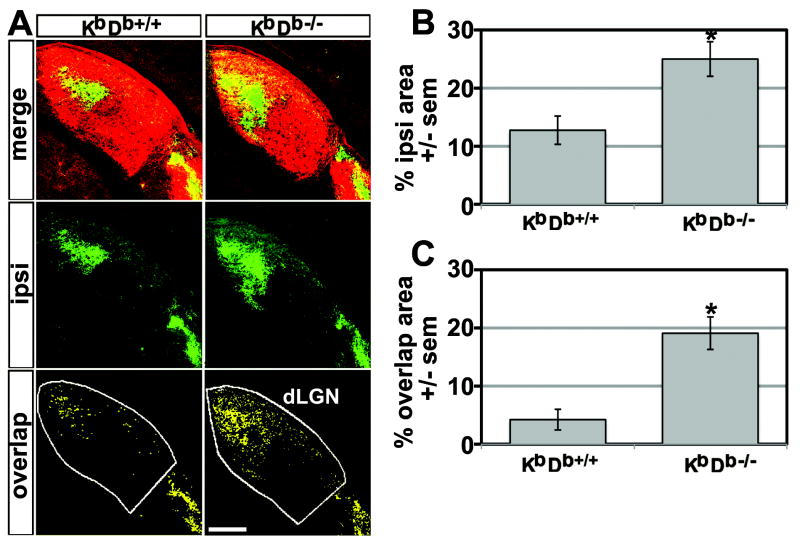
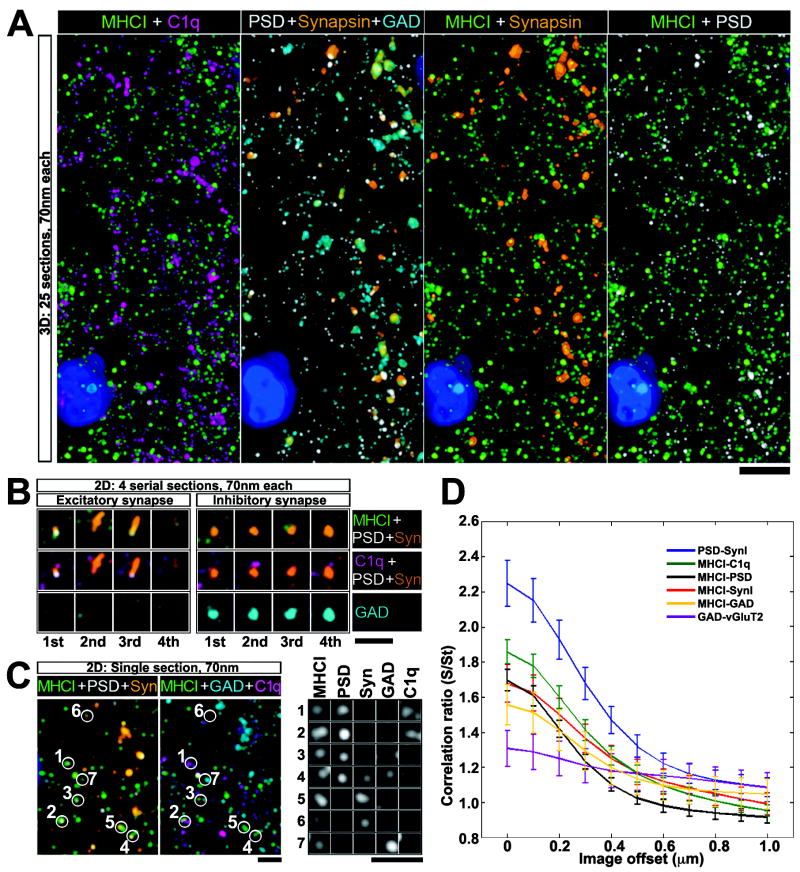
Major histocompatibility complex class I (MHCI) genes were discovered unexpectedly in healthy CNS neurons in a screen for genes regulated by neural activity. In mice lacking just 2 of the 50+ MHCI genes H2-K(b) and H2-D(b), ocular dominance (OD) plasticity is enhanced. Mice lacking PirB, an MHCI receptor, have a similar phenotype. H2-K(b) and H2-D(b) are expressed not only in visual cortex, but also in lateral geniculate nucleus (LGN), where protein localization correlates strongly with synaptic markers and complement protein C1q. In K(b)D(b-/-) mice, developmental refinement of retinogeniculate projections is impaired, similar to C1q(-/-) mice. These phenotypes in K(b)D(b-/-) mice are strikingly similar to those in beta2 m(-/-)TAP1(-/-) mice, which lack cell surface expression of all MHCIs, implying that H2-K(b) and H2-D(b) can account for observed changes in synapse plasticity. H2-K(b) and H2-D(b) ligands, signaling via neuronal MHCI receptors, may enable activity-dependent remodeling of brain circuits during developmental critical periods.
Figures
References
-
- Atwal JK, Pinkston-Gosse J, Syken J, Stawicki S, Wu Y, Shatz C, Tessier-Lavigne M. PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science (New York, N Y. 2008;322:967–970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
