

The morphology of the masticatory apparatus facilitates muscle force production at wide jaw gapes in tree-gouging common marmosets (Callithrix jacchus)
- PMID: 19946083
- PMCID: PMC4075048
- DOI: 10.1242/jeb.029983
The morphology of the masticatory apparatus facilitates muscle force production at wide jaw gapes in tree-gouging common marmosets (Callithrix jacchus)
Abstract
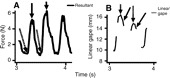
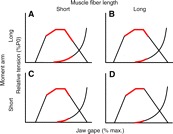
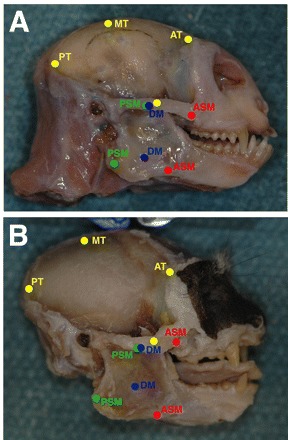
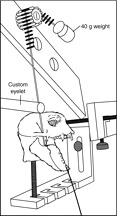
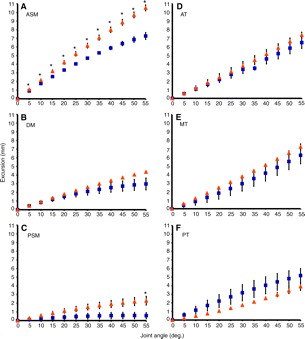
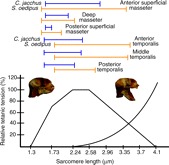
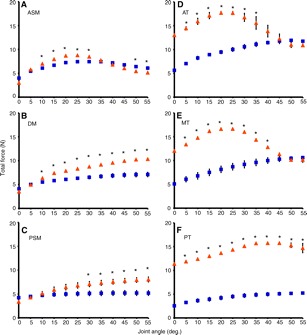
Common marmosets (Callithrix jacchus) generate wide jaw gapes when gouging trees with their anterior teeth to elicit tree exudate flow. Closely related cotton-top tamarins (Saguinus oedipus) do not gouge trees but share similar diets including exudates. Maximizing jaw opening theoretically compromises the bite forces that marmosets can generate during gouging. To investigate how jaw-muscle architecture and craniofacial position impact muscle performance during gouging, we combine skull and jaw-muscle architectural features to model muscle force production across a range of jaw gapes in these two species. We incorporate joint mechanics, resting sarcomere length and muscle architecture estimates from the masseter and temporalis to model muscle excursion, sarcomere length and relative tension as a function of joint angle. Muscle excursion from occlusion to an estimated maximum functional gape of 55 deg. was smaller in all regions of the masseter and temporalis of C. jacchus compared with S. oedipus except the posterior temporalis. As a consequence of reduced muscle excursion distributed over more sarcomeres in series (i.e. longer fibers), sarcomere length operating ranges are smaller in C. jacchus jaw muscles across this range of gapes. This configuration allows C. jacchus to act on a more favorable portion of the length-tension curve at larger gapes and thereby generate relatively greater tension in these muscles compared with S. oedipus. Our results suggest that biting performance during tree gouging in common marmosets is improved by a musculoskeletal configuration that reduces muscle stretch at wide gapes while simultaneously facilitating comparatively large muscle forces at the extremes of jaw opening.
Figures

Similar articles
-
Comparative analysis of masseter fiber architecture in tree-gouging (Callithrix jacchus) and nongouging (Saguinus oedipus) callitrichids.J Morphol. 2004 Sep;261(3):276-85. doi: 10.1002/jmor.10249. J Morphol. 2004. PMID: 15281057
-
The functional correlates of jaw-muscle fiber architecture in tree-gouging and nongouging callitrichid monkeys.Am J Phys Anthropol. 2009 Jul;139(3):353-67. doi: 10.1002/ajpa.20991. Am J Phys Anthropol. 2009. PMID: 19140215
-
Comparative functional analysis of skull morphology of tree-gouging primates.Am J Phys Anthropol. 2003 Feb;120(2):153-70. doi: 10.1002/ajpa.10129. Am J Phys Anthropol. 2003. PMID: 12541333
-
The functional morphology of the anterior masticatory apparatus in tree-gouging marmosets (Cebidae, Primates).J Morphol. 2011 Jul;272(7):833-49. doi: 10.1002/jmor.10951. Epub 2011 Apr 15. J Morphol. 2011. PMID: 21500248
-
Jaw-Muscle Structure and Function in Primates: Insights Into Muscle Performance and Feeding-System Behaviors.Evol Anthropol. 2025 Mar;34(1):e22053. doi: 10.1002/evan.22053. Evol Anthropol. 2025. PMID: 39964129 Free PMC article. Review.
Cited by
-
Comparative Biomechanical Modeling of Metatherian and Placental Saber-Tooths: A Different Kind of Bite for an Extreme Pouched Predator.PLoS One. 2013 Jun 26;8(6):e66888. doi: 10.1371/journal.pone.0066888. Print 2013. PLoS One. 2013. PMID: 23840547 Free PMC article.
-
Comparison of rotator cuff muscle architecture between humans and other selected vertebrate species.J Exp Biol. 2014 Jan 15;217(Pt 2):261-73. doi: 10.1242/jeb.083923. Epub 2013 Sep 26. J Exp Biol. 2014. PMID: 24072803 Free PMC article.
-
Estimating cranial musculoskeletal constraints in theropod dinosaurs.R Soc Open Sci. 2015 Nov 4;2(11):150495. doi: 10.1098/rsos.150495. eCollection 2015 Nov. R Soc Open Sci. 2015. PMID: 26716007 Free PMC article.
-
Bite force is limited by the force-length relationship of skeletal muscle in black carp, Mylopharyngodon piceus.Biol Lett. 2013 Feb 13;9(2):20121181. doi: 10.1098/rsbl.2012.1181. Print 2013 Apr 23. Biol Lett. 2013. PMID: 23407500 Free PMC article.
-
Age-Related Changes in Marmoset (Callithrix jacchus) Feeding Behavior and Physiology: Insights of Masticatory and Swallowing Functions.Am J Primatol. 2025 Aug;87(8):e70070. doi: 10.1002/ajp.70070. Am J Primatol. 2025. PMID: 40855837 Free PMC article.
References
-
- Agrawal K., Lucas P., Bruce I., Prinz J. (1998). Food properties that influence neuromuscular activity during human mastication. J. Dent. Res. 77, 1931-1938 - PubMed
-
- Anapol F., Herring S. (1989). Length-tension relationships of masseter and digastric muscles on miniature swine during ontogeny. J. Exp. Biol. 143, 1-16 - PubMed
-
- Ang K., Lucas P., Tan H. (2006). Incisal orientation and biting efficiency. J. Hum. Evol. 50, 663-672 - PubMed
-
- Blanksma N. G., van Eijden T. M. G. J., van Ruijven L. J., Weijs W. A. (1997). Electromyographic heterogeneity in the human temporalis and masseter muscles during dynamic tasks guided by visual feedback. J. Dent. Res. 76, 542-551 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
