

Stably maintained dendritic spines are associated with lifelong memories
- PMID: 19946265
- PMCID: PMC4724802
- DOI: 10.1038/nature08577
Stably maintained dendritic spines are associated with lifelong memories
Abstract
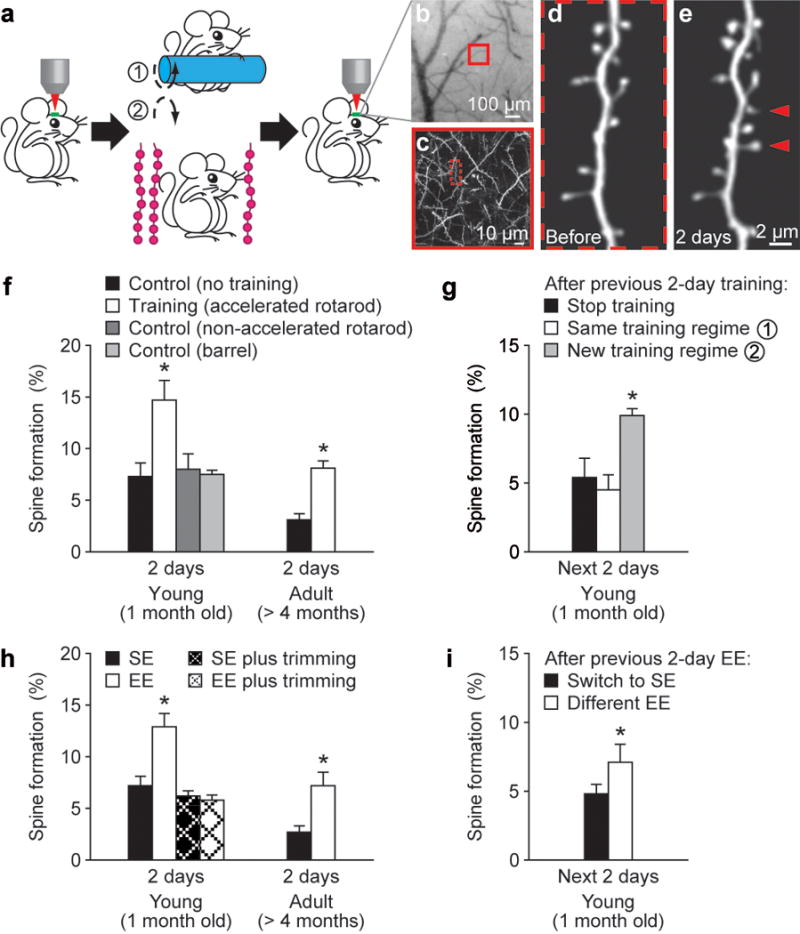
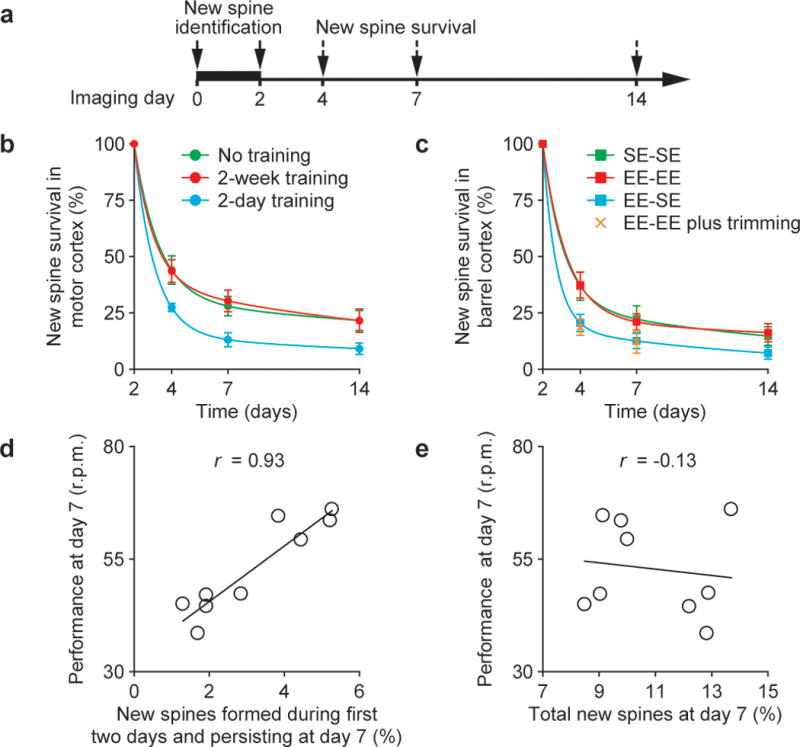
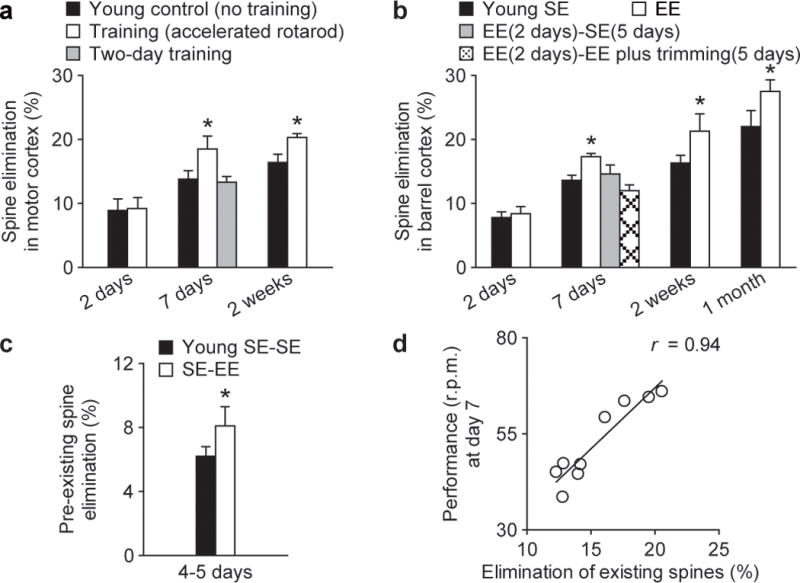
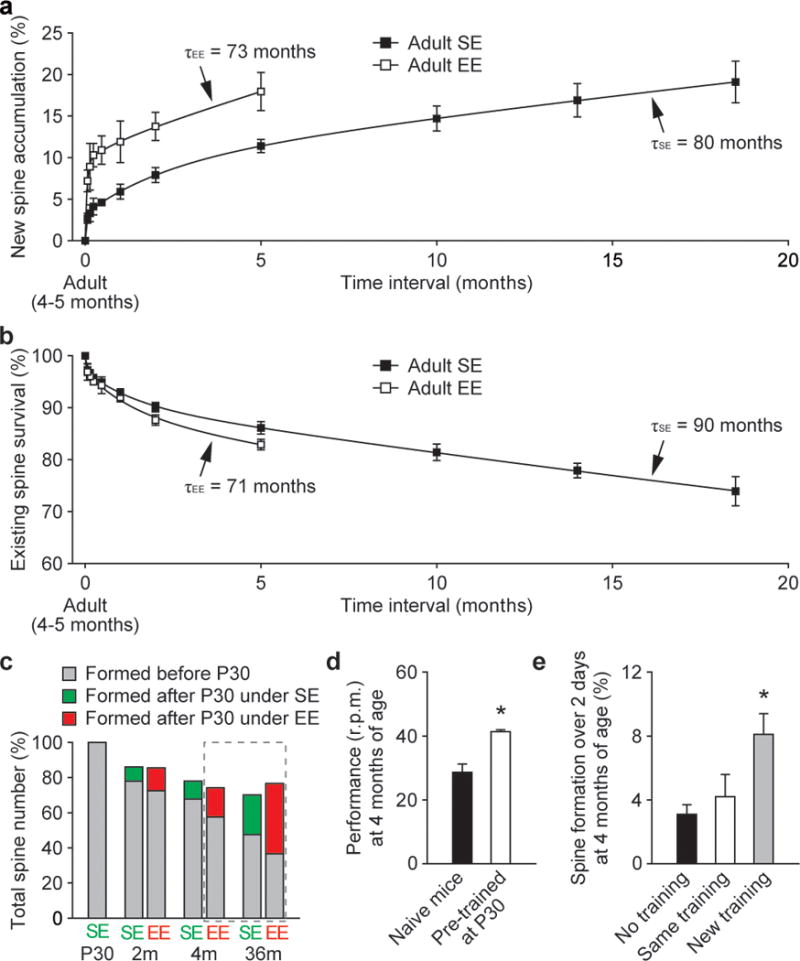
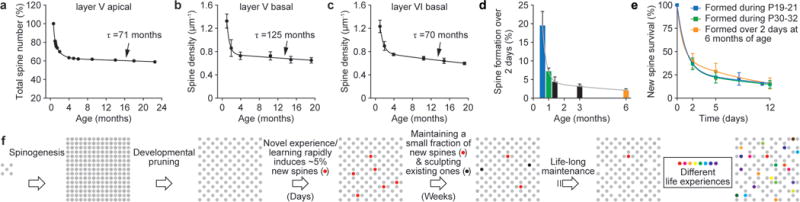
Changes in synaptic connections are considered essential for learning and memory formation. However, it is unknown how neural circuits undergo continuous synaptic changes during learning while maintaining lifelong memories. Here we show, by following postsynaptic dendritic spines over time in the mouse cortex, that learning and novel sensory experience lead to spine formation and elimination by a protracted process. The extent of spine remodelling correlates with behavioural improvement after learning, suggesting a crucial role of synaptic structural plasticity in memory formation. Importantly, a small fraction of new spines induced by novel experience, together with most spines formed early during development and surviving experience-dependent elimination, are preserved and provide a structural basis for memory retention throughout the entire life of an animal. These studies indicate that learning and daily sensory experience leave minute but permanent marks on cortical connections and suggest that lifelong memories are stored in largely stably connected synaptic networks.
Figures
Comment in
-
Neuroscience: New tricks and old spines.Nature. 2009 Dec 17;462(7275):859-61. doi: 10.1038/462859a. Nature. 2009. PMID: 20016588 No abstract available.
References
-
- Bailey CH, Kandel ER. Structural changes accompanying memory storage. Annu Rev Physiol. 1993;55:397–426. - PubMed
-
- Buonomano DV, Merzenich MM. Cortical plasticity: from synapses to maps. Annu Rev Neurosci. 1998;21:149–186. - PubMed
-
- Changeux JP, Danchin A. Selective stabilisation of developing synapses as a mechanism for the specification of neuronal networks. Nature. 1976;264:705–712. - PubMed
-
- Hubel DH, Wiesel TN, LeVay S. Plasticity of ocular dominance columns in monkey striate cortex. Phil Trans R Soc Lond B. 1977;278:377–409. - PubMed
-
- Lichtman JW, Colman H. Synapse elimination and indelible memory. Neuron. 2000;25:269–278. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
