

Intramolecular regulation of phosphorylation status of the circadian clock protein KaiC
- PMID: 19946629
- PMCID: PMC2778140
- DOI: 10.1371/journal.pone.0007509
Intramolecular regulation of phosphorylation status of the circadian clock protein KaiC
Abstract
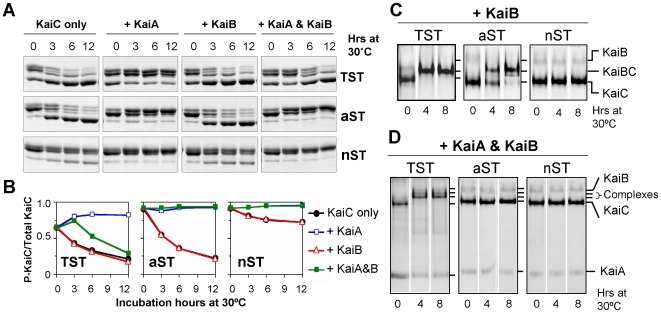
Background: KaiC, a central clock protein in cyanobacteria, undergoes circadian oscillations between hypophosphorylated and hyperphosphorylated forms in vivo and in vitro. Structural analyses of KaiC crystals have identified threonine and serine residues in KaiC at three residues (T426, S431, and T432) as potential sites at which KaiC is phosphorylated; mutation of any of these three sites to alanine abolishes rhythmicity, revealing an essential clock role for each residue separately and for KaiC phosphorylation in general. Mass spectrometry studies confirmed that the S431 and T432 residues are key phosphorylation sites, however, the role of the threonine residue at position 426 was not clear from the mass spectrometry measurements.
Methodology and principal findings: Mutational approaches and biochemical analyses of KaiC support a key role for T426 in control of the KaiC phosphorylation status in vivo and in vitro and demonstrates that alternative amino acids at residue 426 dramatically affect KaiC's properties in vivo and in vitro, especially genetic dominance/recessive relationships, KaiC dephosphorylation, and the formation of complexes of KaiC with KaiA and KaiB. These mutations alter key circadian properties, including period, amplitude, robustness, and temperature compensation. Crystallographic analyses indicate that the T426 site is phosphorylatible under some conditions, and in vitro phosphorylation assays of KaiC demonstrate labile phosphorylation of KaiC when the primary S431 and T432 sites are blocked.
Conclusions and significance: T426 is a crucial site that regulates KaiC phosphorylation status in vivo and in vitro and these studies underscore the importance of KaiC phosphorylation status in the essential cyanobacterial circadian functions. The regulatory roles of these phosphorylation sites--including T426--within KaiC enhance our understanding of the molecular mechanism underlying circadian rhythm generation in cyanobacteria.
Conflict of interest statement
Figures
Similar articles
-
Structures of KaiC circadian clock mutant proteins: a new phosphorylation site at T426 and mechanisms of kinase, ATPase and phosphatase.PLoS One. 2009 Nov 26;4(11):e7529. doi: 10.1371/journal.pone.0007529. PLoS One. 2009. PMID: 19956664 Free PMC article.
-
Dephosphorylation of the core clock protein KaiC in the cyanobacterial KaiABC circadian oscillator proceeds via an ATP synthase mechanism.Biochemistry. 2012 Feb 28;51(8):1547-58. doi: 10.1021/bi201525n. Epub 2012 Feb 13. Biochemistry. 2012. PMID: 22304631 Free PMC article.
-
Identification of key phosphorylation sites in the circadian clock protein KaiC by crystallographic and mutagenetic analyses.Proc Natl Acad Sci U S A. 2004 Sep 21;101(38):13933-8. doi: 10.1073/pnas.0404768101. Epub 2004 Sep 3. Proc Natl Acad Sci U S A. 2004. PMID: 15347809 Free PMC article.
-
A cyanobacterial circadian clock based on the Kai oscillator.Cold Spring Harb Symp Quant Biol. 2007;72:47-55. doi: 10.1101/sqb.2007.72.029. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419262 Review.
-
Diversity of KaiC-based timing systems in marine Cyanobacteria.Mar Genomics. 2014 Apr;14:3-16. doi: 10.1016/j.margen.2013.12.006. Epub 2014 Jan 3. Mar Genomics. 2014. PMID: 24388874 Review.
Cited by
-
Insight into cyanobacterial circadian timing from structural details of the KaiB-KaiC interaction.Proc Natl Acad Sci U S A. 2014 Jan 28;111(4):1379-84. doi: 10.1073/pnas.1314326111. Epub 2014 Jan 13. Proc Natl Acad Sci U S A. 2014. PMID: 24474762 Free PMC article.
-
Minimal tool set for a prokaryotic circadian clock.BMC Evol Biol. 2017 Jul 21;17(1):169. doi: 10.1186/s12862-017-0999-7. BMC Evol Biol. 2017. PMID: 28732467 Free PMC article.
-
The itty-bitty time machine genetics of the cyanobacterial circadian clock.Adv Genet. 2011;74:13-53. doi: 10.1016/B978-0-12-387690-4.00002-7. Adv Genet. 2011. PMID: 21924974 Free PMC article. Review.
-
KaiC intersubunit communication facilitates robustness of circadian rhythms in cyanobacteria.Nat Commun. 2013;4:2897. doi: 10.1038/ncomms3897. Nat Commun. 2013. PMID: 24305644 Free PMC article.
-
Combined SAXS/EM based models of the S. elongatus post-translational circadian oscillator and its interactions with the output His-kinase SasA.PLoS One. 2011;6(8):e23697. doi: 10.1371/journal.pone.0023697. Epub 2011 Aug 24. PLoS One. 2011. PMID: 21887298 Free PMC article.
References
-
- Ditty JL, Mackey SR, Johnson CH. Springer: Verlag Berlin Heidelberg; 2009. Bacterial Circadian Programs.333
-
- Ishiura M, Kutsuna S, Aoki S, Iwasaki H, Andersson CR, et al. Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science. 1998;281:1519–1523. - PubMed
-
- Nakajima M, Imai K, Ito H, Nishiwaki T, Murayama Y, et al. Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro. Science. 2005;308:414–415. - PubMed
-
- Tomita J, Nakajima M, Kondo T, Iwasaki H. No transcription-translation feedback in circadian rhythm of KaiC phosphorylation. Science. 2005;307:251–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
