

Zebrafish con/disp1 reveals multiple spatiotemporal requirements for Hedgehog-signaling in craniofacial development
- PMID: 19948063
- PMCID: PMC2791760
- DOI: 10.1186/1471-213X-9-59
Zebrafish con/disp1 reveals multiple spatiotemporal requirements for Hedgehog-signaling in craniofacial development
Abstract
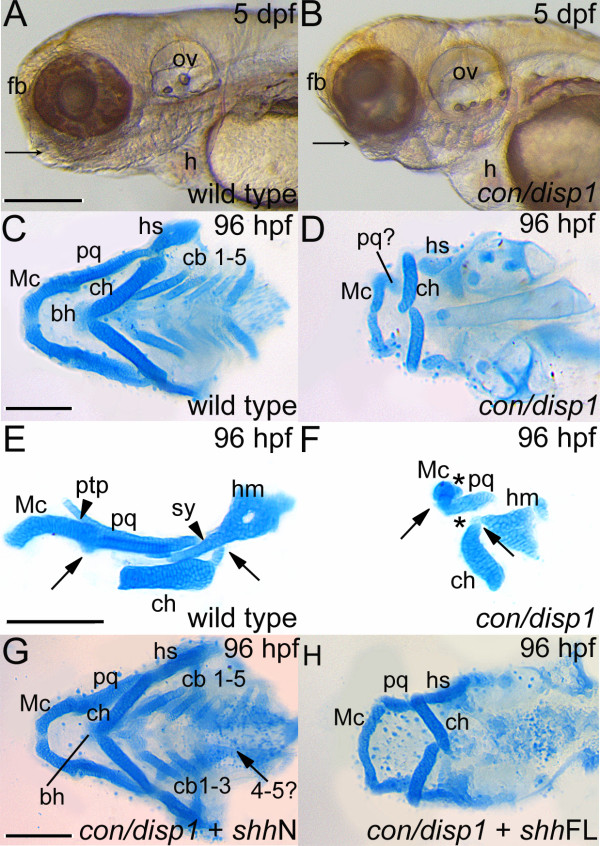
Background: The vertebrate head skeleton is derived largely from cranial neural crest cells (CNCC). Genetic studies in zebrafish and mice have established that the Hedgehog (Hh)-signaling pathway plays a critical role in craniofacial development, partly due to the pathway's role in CNCC development. Disruption of the Hh-signaling pathway in humans can lead to the spectral disorder of Holoprosencephaly (HPE), which is often characterized by a variety of craniofacial defects including midline facial clefting and cyclopia 12. Previous work has uncovered a role for Hh-signaling in zebrafish dorsal neurocranium patterning and chondrogenesis, however Hh-signaling mutants have not been described with respect to the ventral pharyngeal arch (PA) skeleton. Lipid-modified Hh-ligands require the transmembrane-spanning receptor Dispatched 1 (Disp1) for proper secretion from Hh-synthesizing cells to the extracellular field where they act on target cells. Here we study chameleon mutants, lacking a functional disp1(con/disp1).
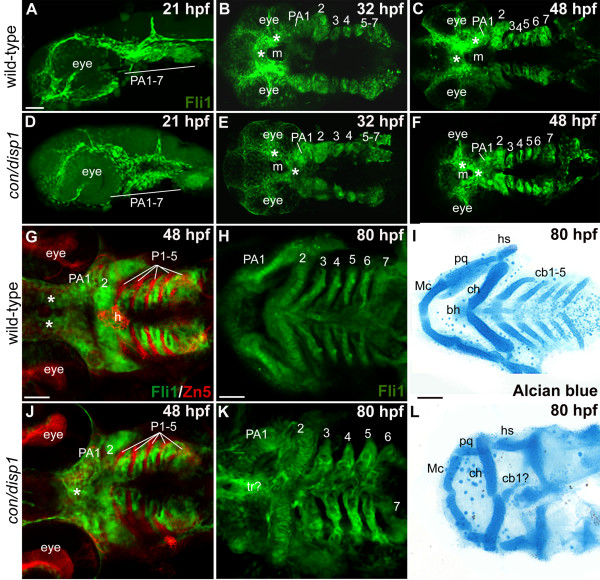
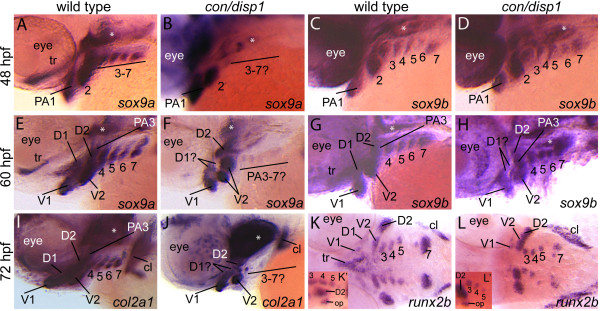
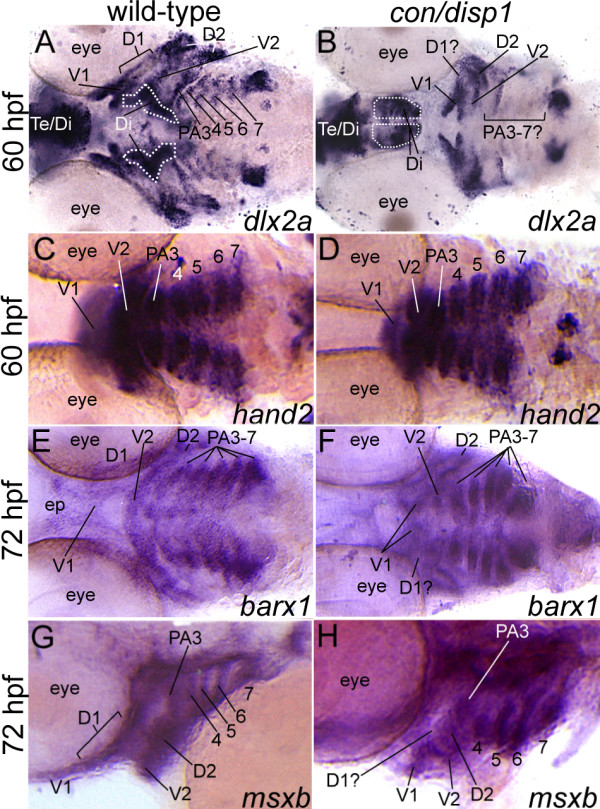
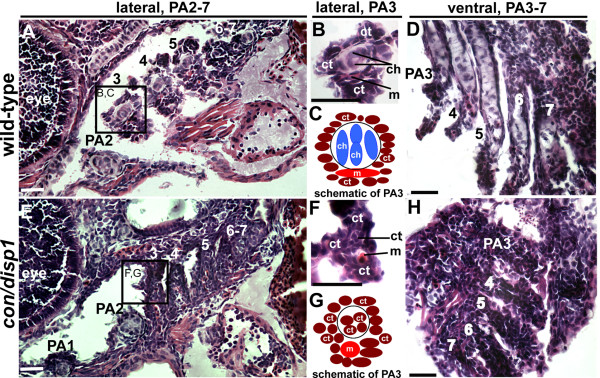
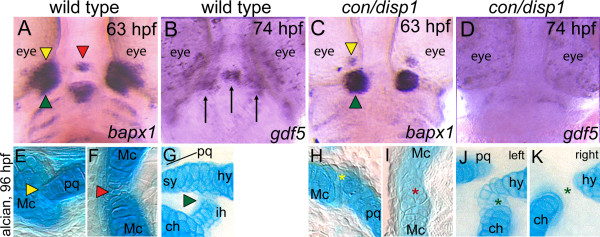
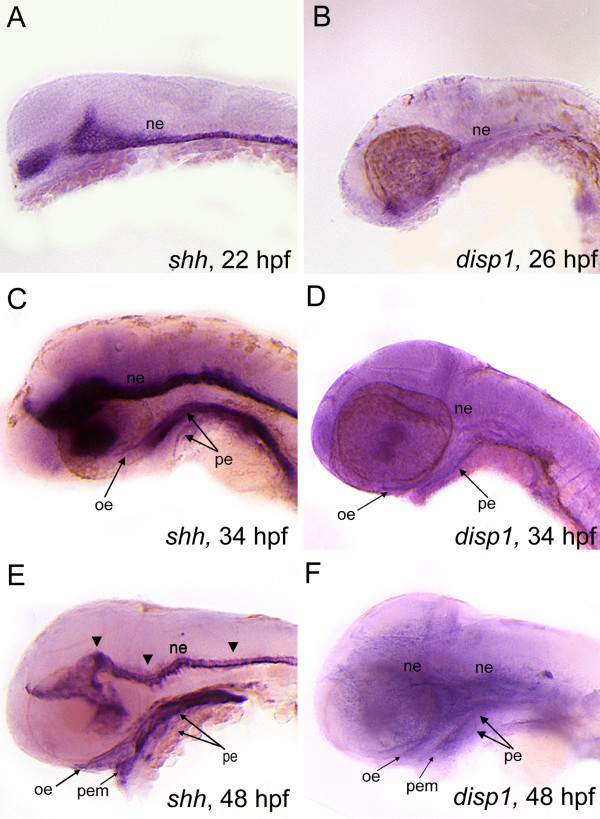
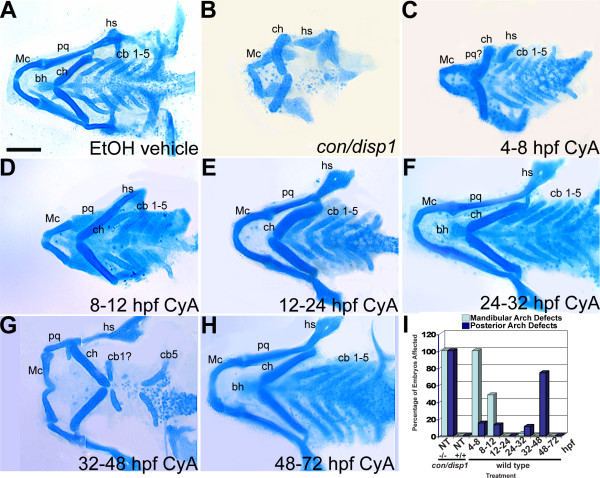
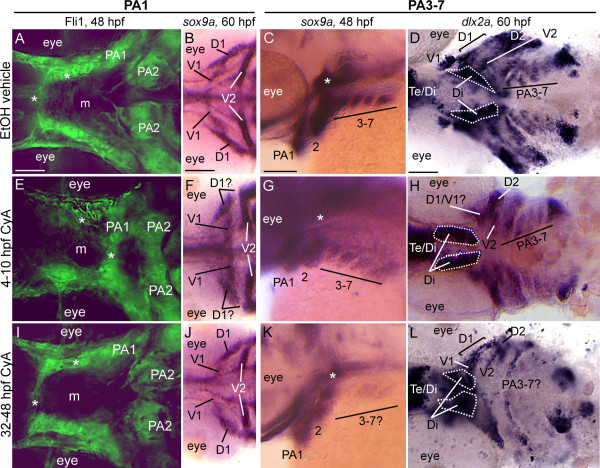
Results: con/disp1 mutants display reduced and dysmorphic mandibular and hyoid arch cartilages and lack all ceratobranchial cartilage elements. CNCC specification and migration into the PA primorida occurs normally in con/disp1 mutants, however disp1 is necessary for post-migratory CNCC patterning and differentiation. We show that disp1 is required for post-migratory CNCC to become properly patterned within the first arch, while the gene is dispensable for CNCC condensation and patterning in more posterior arches. Upon residing in well-formed pharyngeal epithelium, neural crest condensations in the posterior PA fail to maintain expression of two transcription factors essential for chondrogenesis, sox9a and dlx2a, yet continue to robustly express other neural crest markers. Histology reveals that posterior arch residing-CNCC differentiate into fibrous-connective tissue, rather than becoming chondrocytes. Treatments with Cyclopamine, to inhibit Hh-signaling at different developmental stages, show that Hh-signaling is required during gastrulation for normal patterning of CNCC in the first PA, and then during the late pharyngula stage, to promote CNCC chondrogenesis within the posterior arches. Further, loss of disp1 disrupted normal expression of bapx1 and gdf5, markers of jaw joint patterning, thus resulting in jaw joint defects in con/disp1 mutant animals.
Conclusion: This study reveals novel requirements for Hh-signaling in the zebrafish PA skeleton and highlights the functional diversity and differential sensitivity of craniofacial tissues to Hh-signaling throughout the face, a finding that may help to explain the spectrum of human facial phenotypes characteristic of HPE.
Figures
Similar articles
-
Hh signaling regulates patterning and morphogenesis of the pharyngeal arch-derived skeleton.Dev Biol. 2012 Sep 1;369(1):65-75. doi: 10.1016/j.ydbio.2012.05.032. Epub 2012 Jun 16. Dev Biol. 2012. PMID: 22709972 Free PMC article.
-
Hedgehog signaling is required for cranial neural crest morphogenesis and chondrogenesis at the midline in the zebrafish skull.Development. 2005 Sep;132(17):3977-88. doi: 10.1242/dev.01943. Epub 2005 Jul 27. Development. 2005. PMID: 16049113
-
Early Hedgehog signaling from neural to oral epithelium organizes anterior craniofacial development.Development. 2006 Mar;133(6):1069-77. doi: 10.1242/dev.02281. Epub 2006 Feb 15. Development. 2006. PMID: 16481351
-
Evolvability of the vertebrate craniofacial skeleton.Semin Cell Dev Biol. 2019 Jul;91:13-22. doi: 10.1016/j.semcdb.2017.12.004. Epub 2017 Dec 13. Semin Cell Dev Biol. 2019. PMID: 29248471 Free PMC article. Review.
-
Molecular dissection of craniofacial development using zebrafish.Crit Rev Oral Biol Med. 2002;13(4):308-22. doi: 10.1177/154411130201300402. Crit Rev Oral Biol Med. 2002. PMID: 12191958 Review.
Cited by
-
Reply to Huysseune and Witten: Oral tooth bud formation in zebrafish.Proc Natl Acad Sci U S A. 2024 Aug 27;121(35):e2413644121. doi: 10.1073/pnas.2413644121. Epub 2024 Aug 16. Proc Natl Acad Sci U S A. 2024. PMID: 39150774 Free PMC article. No abstract available.
-
Visualization of Gli activity in craniofacial tissues of hedgehog-pathway reporter transgenic zebrafish.PLoS One. 2010 Dec 21;5(12):e14396. doi: 10.1371/journal.pone.0014396. PLoS One. 2010. PMID: 21203590 Free PMC article.
-
Pdlim7 is required for maintenance of the mesenchymal/epidermal Fgf signaling feedback loop during zebrafish pectoral fin development.BMC Dev Biol. 2010 Oct 15;10:104. doi: 10.1186/1471-213X-10-104. BMC Dev Biol. 2010. PMID: 20950450 Free PMC article.
-
Pharmacological activation of the Sonic hedgehog pathway with a Smoothened small molecule agonist ameliorates the severity of alcohol-induced morphological and behavioral birth defects in a zebrafish model of fetal alcohol spectrum disorder.J Neurosci Res. 2022 Aug;100(8):1585-1601. doi: 10.1002/jnr.25008. Epub 2022 Jan 11. J Neurosci Res. 2022. PMID: 35014067 Free PMC article.
-
A new mechanistic scenario for the origin and evolution of vertebrate cartilage.PLoS One. 2011;6(7):e22474. doi: 10.1371/journal.pone.0022474. Epub 2011 Jul 22. PLoS One. 2011. PMID: 21799866 Free PMC article.
References
-
- Couly G, Creuzet S, Bennaceur S, Vincent C, Le Douarin NM. Interactions between Hox-negative cephalic neural crest cells and the foregut endoderm in patterning the facial skeleton in the vertebrate head. Development. 2002;129:1061–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
