

Role of Scrib and Dlg in anterior-posterior patterning of the follicular epithelium during Drosophila oogenesis
- PMID: 19948068
- PMCID: PMC2810132
- DOI: 10.1186/1471-213X-9-60
Role of Scrib and Dlg in anterior-posterior patterning of the follicular epithelium during Drosophila oogenesis
Abstract
Background: Proper patterning of the follicle cell epithelium over the egg chamber is essential for the Drosophila egg development. Differentiation of the epithelium into several distinct cell types along the anterior-posterior axis requires coordinated activities of multiple signaling pathways. Previously, we reported that lethal(2)giant larvae (lgl), a Drosophila tumor suppressor gene, is required in the follicle cells for the posterior follicle cell (PFC) fate induction at mid-oogenesis. Here we explore the role of another two tumor suppressor genes, scribble (scrib) and discs large (dlg), in the epithelial patterning.
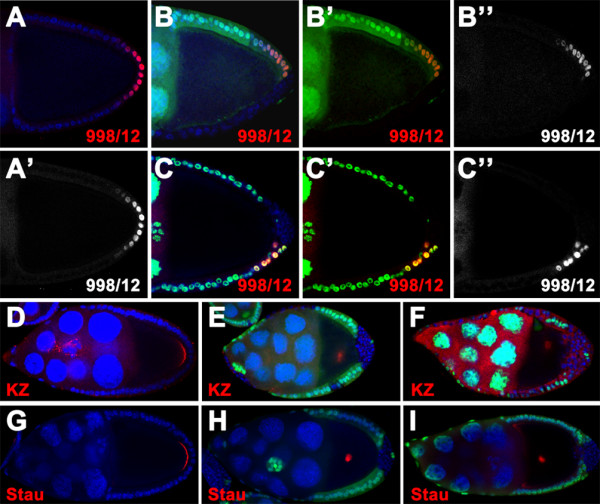
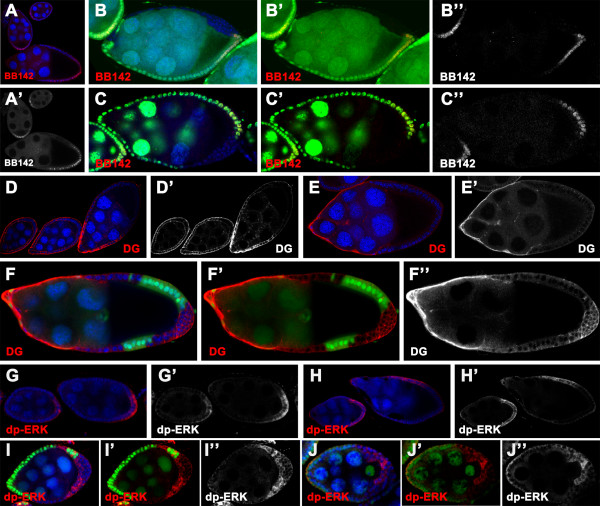
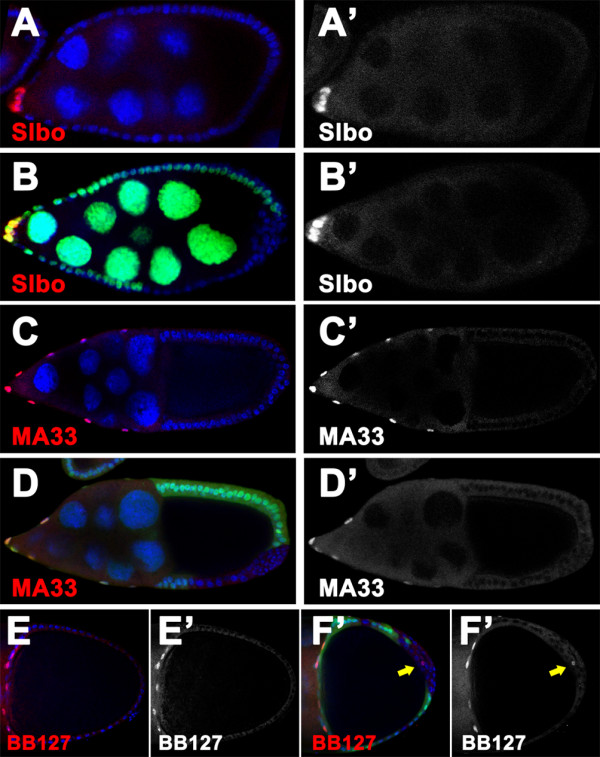
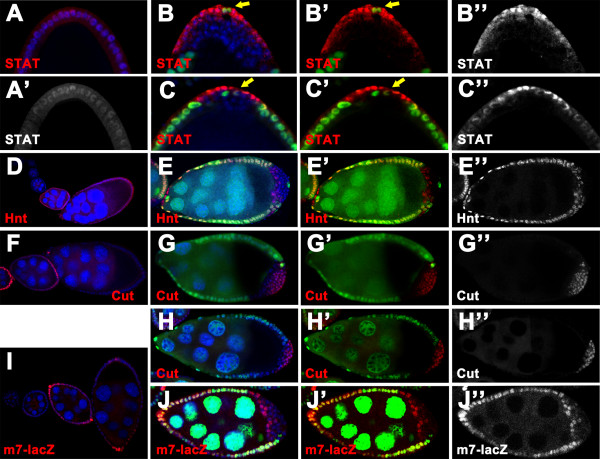
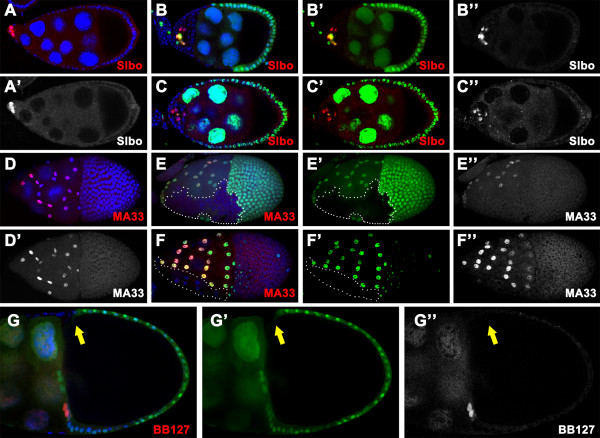
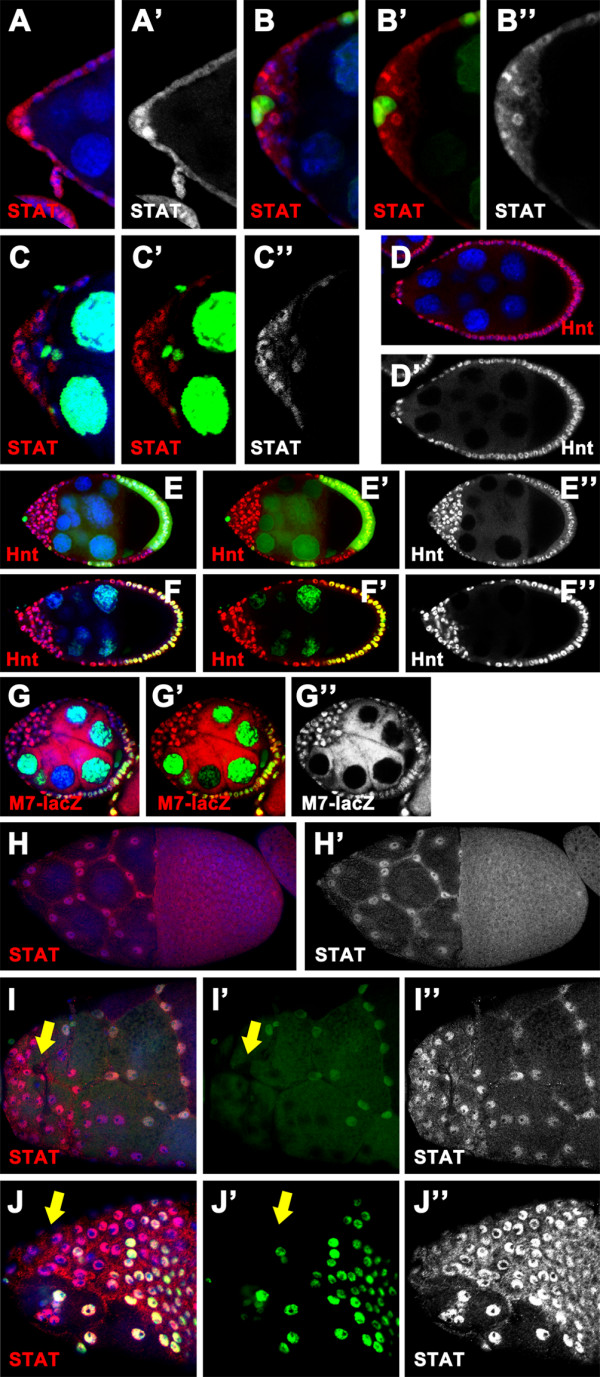
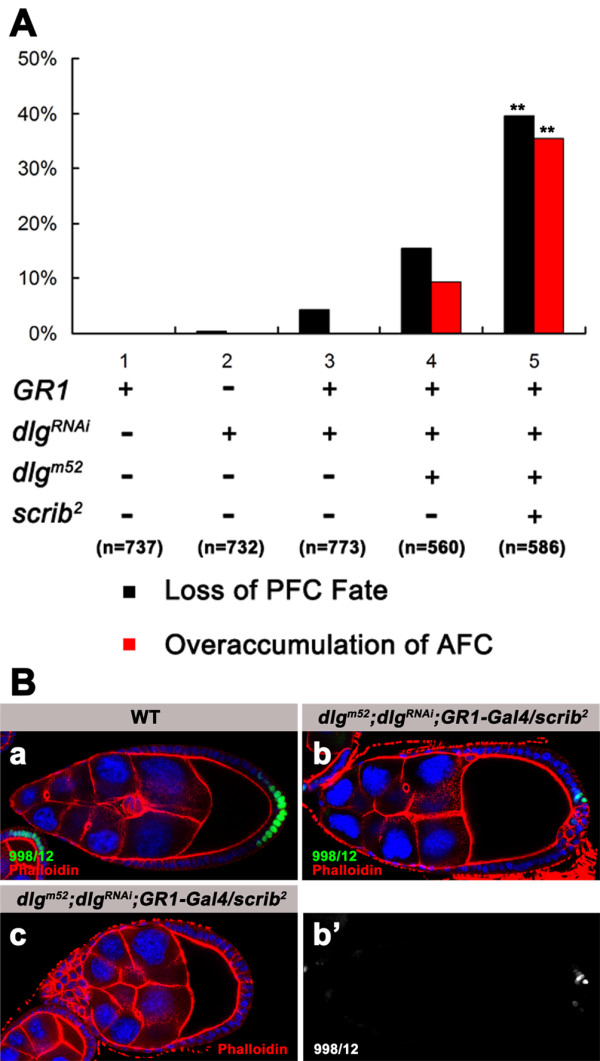
Results: We found that removal of scrib or dlg function from the follicle cells at posterior terminal of the egg chamber causes a complete loss of the PFC fate. Aberrant specification and differentiation of the PFCs in the mosaic clones can be ascribed to defects in coordinated activation of the EGFR, JAK and Notch signaling pathways in the multilayered cells. Meanwhile, the clonal analysis revealed that loss-of-function mutations in scrib/dlg at the anterior domains result in a partially penetrant phenotype of defective induction of the stretched and centripetal cell fate, whereas specification of the border cell fate can still occur in the most anterior region of the mutant clones. Further, we showed that scrib genetically interacts with dlg in regulating posterior patterning of the epithelium.
Conclusion: In this study we provide evidence that scrib and dlg function differentially in anterior and posterior patterning of the follicular epithelium at oogenesis. Further genetic analysis indicates that scrib and dlg act in a common pathway to regulate PFC fate induction. This study may open another window for elucidating role of scrib/dlg in controlling epithelial polarity and cell proliferation during development.
Figures
Similar articles
-
Requirements of Lgl in cell differentiation and motility during Drosophila ovarian follicular epithelium morphogenesis.Fly (Austin). 2011 Apr-Jun;5(2):81-7. doi: 10.4161/fly.5.2.14436. Epub 2011 Apr 1. Fly (Austin). 2011. PMID: 21245664
-
Lgl cortical dynamics are independent of binding to the Scrib-Dlg complex but require Dlg-dependent restriction of aPKC.Development. 2020 Aug 7;147(15):dev186593. doi: 10.1242/dev.186593. Development. 2020. PMID: 32665243
-
Distinct activities of Scrib module proteins organize epithelial polarity.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11531-11540. doi: 10.1073/pnas.1918462117. Epub 2020 May 15. Proc Natl Acad Sci U S A. 2020. PMID: 32414916 Free PMC article.
-
Dlg, Scribble and Lgl in cell polarity, cell proliferation and cancer.Bioessays. 2003 Jun;25(6):542-53. doi: 10.1002/bies.10286. Bioessays. 2003. PMID: 12766944 Review.
-
Signaling between somatic follicle cells and the germline patterns the egg and embryo of Drosophila.Curr Top Dev Biol. 2020;140:55-86. doi: 10.1016/bs.ctdb.2019.10.004. Epub 2019 Nov 19. Curr Top Dev Biol. 2020. PMID: 32591083 Review.
Cited by
-
Notch Transduction in Non-Small Cell Lung Cancer.Int J Mol Sci. 2020 Aug 8;21(16):5691. doi: 10.3390/ijms21165691. Int J Mol Sci. 2020. PMID: 32784481 Free PMC article. Review.
-
Characterization of the Src/Abl hybrid kinase SmTK6 of Schistosoma mansoni.J Biol Chem. 2011 Dec 9;286(49):42325-42336. doi: 10.1074/jbc.M110.210336. Epub 2011 Oct 19. J Biol Chem. 2011. PMID: 22013071 Free PMC article.
-
Synaptic components are required for glioblastoma progression in Drosophila.PLoS Genet. 2022 Jul 25;18(7):e1010329. doi: 10.1371/journal.pgen.1010329. eCollection 2022 Jul. PLoS Genet. 2022. PMID: 35877760 Free PMC article.
-
At the crossroads of differentiation and proliferation: precise control of cell-cycle changes by multiple signaling pathways in Drosophila follicle cells.Bioessays. 2011 Feb;33(2):124-34. doi: 10.1002/bies.201000089. Bioessays. 2011. PMID: 21154780 Free PMC article. Review.
-
ArfGAP1 regulates the endosomal sorting of guidance receptors to promote directed collective cell migration in vivo.iScience. 2023 Jul 25;26(8):107467. doi: 10.1016/j.isci.2023.107467. eCollection 2023 Aug 18. iScience. 2023. PMID: 37599820 Free PMC article.
References
-
- Gonzalez-Reyes A, St Johnston D. Patterning of the follicle cell epithelium along the anterior-posterior axis during Drosophila oogenesis. Development. 1998;125:2837–2846. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
