

Helix/coil nucleation: a local response to global demands
- PMID: 19948130
- PMCID: PMC2784565
- DOI: 10.1016/j.bpj.2009.09.013
Helix/coil nucleation: a local response to global demands
Abstract
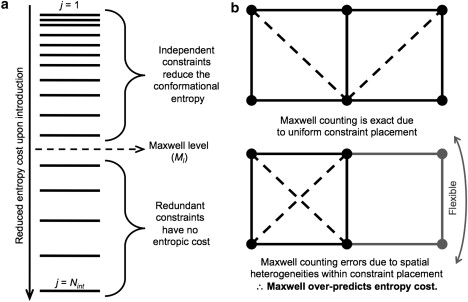
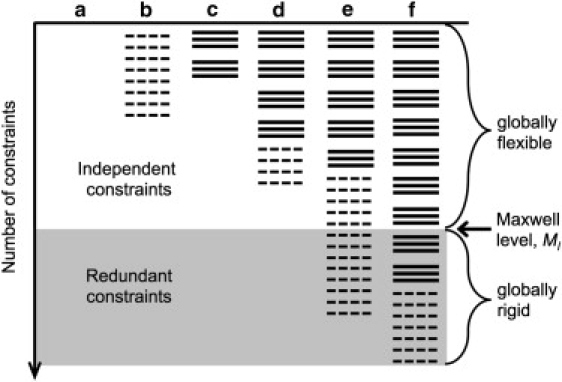
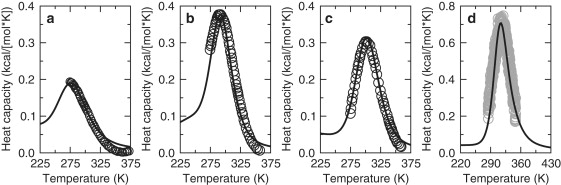
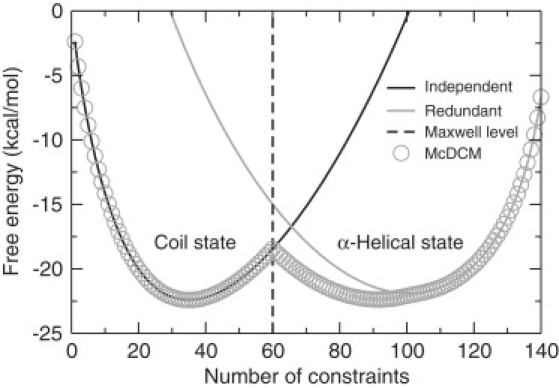
A complete description of protein structure and function must include a proper treatment of mechanisms that lead to cooperativity. The helix/coil transition serves as a simple example of a cooperative folding process, commonly described by a nucleation-propagation mechanism. The prevalent view is that coil structure must first form a short segment of helix in a localized region despite paying a free energy cost (nucleation). Afterward, helical structure propagates outward from the nucleation site. Both processes entail enthalpy-entropy compensation that derives from the loss in conformational entropy on helix formation with concomitant gain in favorable interactions. Nucleation-propagation models inherently assume that cooperativity arises from a sequential series of local events. An alternative distance constraint model asserts there is a direct link between available degrees of freedom and cooperativity through the nonadditivity in conformational entropy. That is, helix nucleation is a concerted manifestation of rigidity propagating through atomic structure. The link between network rigidity and nonadditivity of conformational entropy is shown in this study by solving the distance constraint model using a simple global constraint counting approximation. Cooperativity arises from competition between excess and deficiency in available degrees of freedom in the coil and helix states respectively.
Figures
Similar articles
-
Nonadditivity in the alpha-helix to coil transition.Biopolymers. 2011 Apr;95(4):240-53. doi: 10.1002/bip.21572. Epub 2010 Dec 23. Biopolymers. 2011. PMID: 21280020 Free PMC article.
-
Nonadditivity in conformational entropy upon molecular rigidification reveals a universal mechanism affecting folding cooperativity.Biophys J. 2011 Feb 16;100(4):1129-38. doi: 10.1016/j.bpj.2011.01.027. Biophys J. 2011. PMID: 21320459 Free PMC article.
-
Network rigidity at finite temperature: relationships between thermodynamic stability, the nonadditivity of entropy, and cooperativity in molecular systems.Phys Rev E Stat Nonlin Soft Matter Phys. 2003 Dec;68(6 Pt 1):061109. doi: 10.1103/PhysRevE.68.061109. Epub 2003 Dec 31. Phys Rev E Stat Nonlin Soft Matter Phys. 2003. PMID: 14754182 Free PMC article.
-
Free energy determinants of secondary structure formation: I. alpha-Helices.J Mol Biol. 1995 Sep 22;252(3):351-65. doi: 10.1006/jmbi.1995.0502. J Mol Biol. 1995. PMID: 7563056
-
Helix-coil transition theories. Are they correct?Acta Biochim Pol. 1997;44(3):423-32. Acta Biochim Pol. 1997. PMID: 9511955 Review.
Cited by
-
A virtual pebble game to ensemble average graph rigidity.Algorithms Mol Biol. 2015 Mar 18;10:11. doi: 10.1186/s13015-015-0039-3. eCollection 2015. Algorithms Mol Biol. 2015. PMID: 25904973 Free PMC article.
-
Variations within class-A β-lactamase physiochemical properties reflect evolutionary and environmental patterns, but not antibiotic specificity.PLoS Comput Biol. 2013;9(7):e1003155. doi: 10.1371/journal.pcbi.1003155. Epub 2013 Jul 18. PLoS Comput Biol. 2013. PMID: 23874193 Free PMC article.
-
AIM for Allostery: Using the Ising Model to Understand Information Processing and Transmission in Allosteric Biomolecular Systems.Entropy (Basel). 2015 May;17(5):2895-2918. doi: 10.3390/e17052895. Epub 2015 May 7. Entropy (Basel). 2015. PMID: 26594108 Free PMC article.
-
Nonadditivity in the alpha-helix to coil transition.Biopolymers. 2011 Apr;95(4):240-53. doi: 10.1002/bip.21572. Epub 2010 Dec 23. Biopolymers. 2011. PMID: 21280020 Free PMC article.
-
Redistribution of flexibility in stabilizing antibody fragment mutants follows Le Châtelier's principle.PLoS One. 2014 Mar 26;9(3):e92870. doi: 10.1371/journal.pone.0092870. eCollection 2014. PLoS One. 2014. PMID: 24671209 Free PMC article.
References
-
- IUPAC . Blackwell Scientific Publications; Oxford: 1997. Compendium of Chemical Terminology.
-
- Miranker A.D., Dobson C.M. Collapse and cooperativity in protein folding. Curr. Opin. Struct. Biol. 1996;6:31–42. - PubMed
-
- Krusek J. Allostery and cooperativity in the interaction of drugs with ionic channel receptors. Physiol. Res. 2004;53:569–579. - PubMed
-
- Whitty A. Cooperativity and biological complexity. Nat. Chem. Biol. 2008;4:435–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
