

The ubiquitin landscape at DNA double-strand breaks
- PMID: 19948475
- PMCID: PMC2779242
- DOI: 10.1083/jcb.200908074
The ubiquitin landscape at DNA double-strand breaks
Abstract
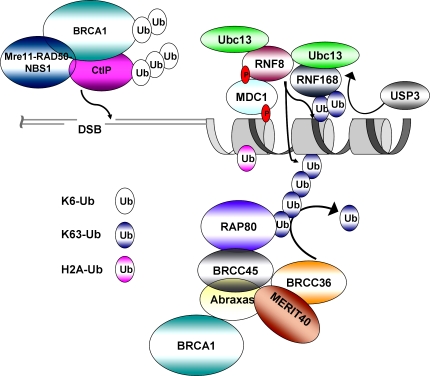
The intimate relationship between DNA double-strand break (DSB) repair and cancer susceptibility has sparked profound interest in how transactions on DNA and chromatin surrounding DNA damage influence genome integrity. Recent evidence implicates a substantial commitment of the cellular DNA damage response machinery to the synthesis, recognition, and hydrolysis of ubiquitin chains at DNA damage sites. In this review, we propose that, in order to accommodate parallel processes involved in DSB repair and checkpoint signaling, DSB-associated ubiquitin structures must be nonuniform, using different linkages for distinct functional outputs. We highlight recent advances in the study of nondegradative ubiquitin signaling at DSBs, and discuss how recognition of different ubiquitin structures may influence DNA damage responses.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
