

Centrosomal Aki1 and cohesin function in separase-regulated centriole disengagement
- PMID: 19948489
- PMCID: PMC2806580
- DOI: 10.1083/jcb.200906019
Centrosomal Aki1 and cohesin function in separase-regulated centriole disengagement
Abstract
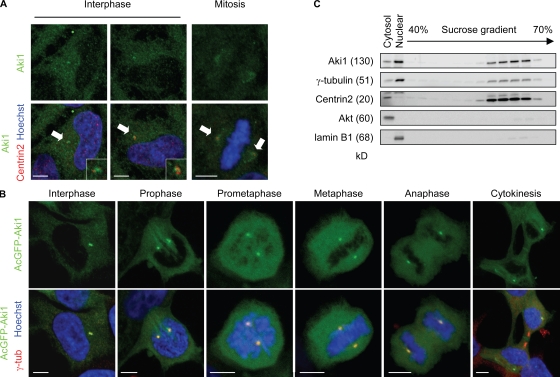
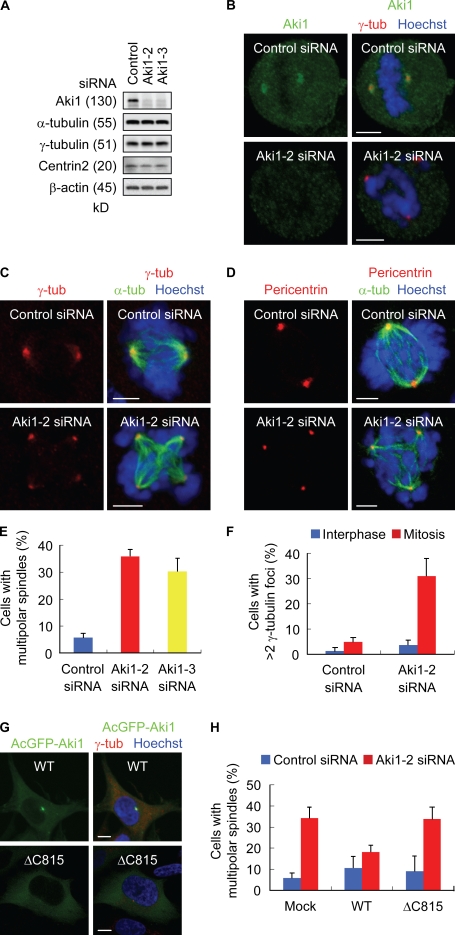
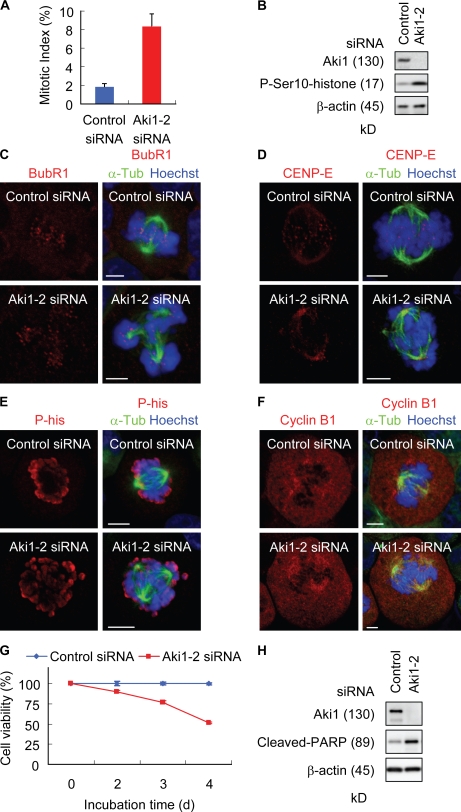
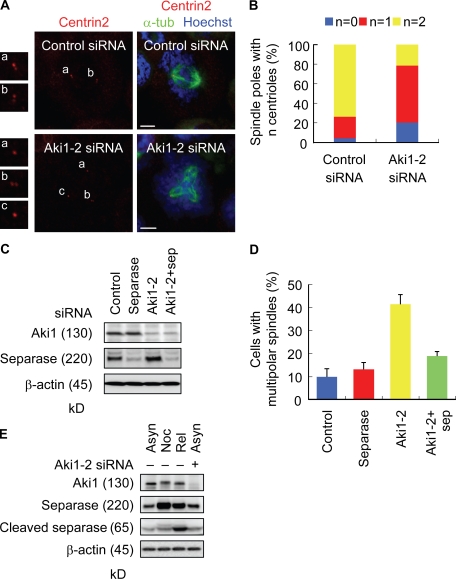
Sister chromatid separation at anaphase is triggered by cleavage of the cohesin subunit Scc1, which is mediated by separase. Centriole disengagement also requires separase. This dual role of separase permits concurrent control of these events for accurate metaphase to anaphase transition. Although the molecular mechanism underlying sister chromatid cohesion has been clarified, that of centriole cohesion is poorly understood. In this study, we show that Akt kinase-interacting protein 1 (Aki1) localizes to centrosomes and regulates centriole cohesion. Aki1 depletion causes formation of multipolar spindles accompanied by centriole splitting, which is separase dependent. We also show that cohesin subunits localize to centrosomes and that centrosomal Scc1 is cleaved by separase coincidentally with chromatin Scc1, suggesting a role of Scc1 as a connector of centrioles as well as sister chromatids. Interestingly, Scc1 depletion strongly induces centriole splitting. Furthermore, Aki1 interacts with cohesin in centrosomes, and this interaction is required for centriole cohesion. We demonstrate that centrosome-associated Aki1 and cohesin play pivotal roles in preventing premature cleavage in centriole cohesion.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
