

Mitochondrial C1-tetrahydrofolate synthase (MTHFD1L) supports the flow of mitochondrial one-carbon units into the methyl cycle in embryos
- PMID: 19948730
- PMCID: PMC2836066
- DOI: 10.1074/jbc.M109.079855
Mitochondrial C1-tetrahydrofolate synthase (MTHFD1L) supports the flow of mitochondrial one-carbon units into the methyl cycle in embryos
Abstract
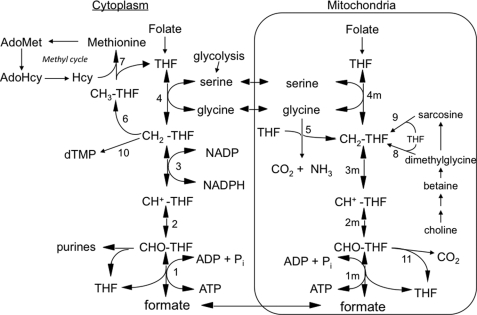
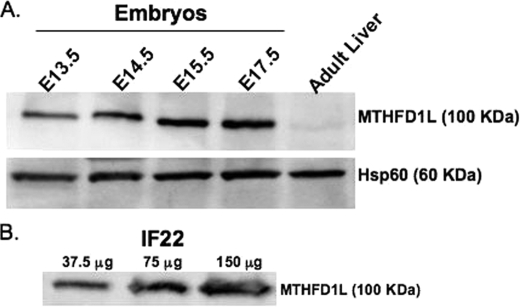
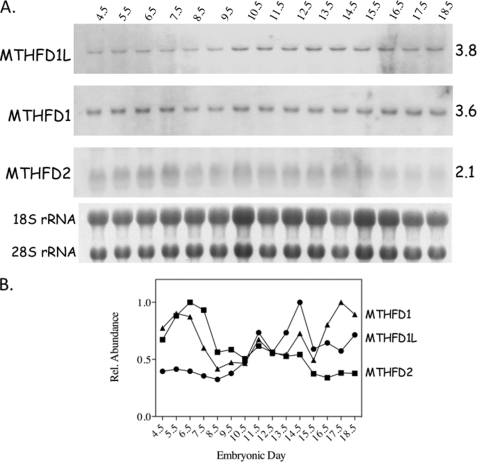
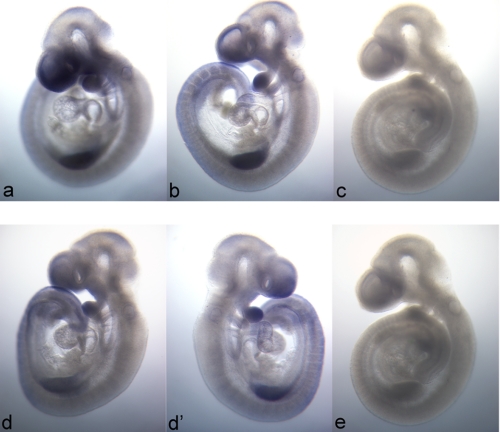
Mitochondrial folate-dependent one-carbon (1-C) metabolism converts 1-C donors such as serine and glycine to formate, which is exported and incorporated into the cytoplasmic tetrahydrofolate (THF) 1-C pool. Developing embryos depend on this mitochondrial pathway to provide 1-C units for cytoplasmic process such as de novo purine biosynthesis and the methyl cycle. This pathway is composed of sequential methylene-THF dehydrogenase, methenyl-THF cyclohydrolase, and 10-formyl-THF synthetase activities. In embryonic mitochondria, the bifunctional MTHFD2 enzyme catalyzes the dehydrogenase and cyclohydrolase reactions, but the enzyme responsible for the mitochondrial synthetase reaction has not been identified in embryos. A monofunctional 10-formyl-THF synthetase (MTHFD1L gene product) functions in adult mitochondria and is a likely candidate for the embryonic activity. Here we show that the MTHFD1L enzyme is present in mitochondria from normal embryonic tissues and embryonic fibroblast cell lines, and embryonic mitochondria possess the ability to synthesize formate from glycine. The MTHFD1L transcript was detected at all stages of mouse embryogenesis examined. In situ hybridizations showed that MTHFD1L was expressed ubiquitously throughout the embryo but with localized regions of higher expression. The spatial pattern of MTHFD1L expression was virtually indistinguishable from that of MTHFD2 and MTHFD1 (cytoplasmic C(1)-THF synthase) in embryonic day 9.5 mouse embryos, suggesting coordinated regulation. Finally, we show using stable isotope labeling that in an embryonic mouse cell line, greater than 75% of 1-C units entering the cytoplasmic methyl cycle are mitochondrially derived. Thus, a complete pathway of enzymes for supplying 1-C units from the mitochondria to the methyl cycle in embryonic tissues is established.
Figures

References
-
- Appling D. R. (1991) FASEB J. 5, 2645–2651 - PubMed
-
- Woeller C. F., Anderson D. D., Szebenyi D. M., Stover P. J. (2007) J. Biol. Chem. 282, 17623–17631 - PubMed
-
- Davis S. R., Stacpoole P. W., Williamson J., Kick L. S., Quinlivan E. P., Coats B. S., Shane B., Bailey L. B., Gregory J. F., 3rd (2004) Am. J. Physiol. Endocrinol. Metab. 286, E272–E279 - PubMed
-
- Fu T. F., Rife J. P., Schirch V. (2001) Arch. Biochem. Biophys. 393, 42–50 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
