

Control of cell proliferation, organ growth, and DNA damage response operate independently of dephosphorylation of the Arabidopsis Cdk1 homolog CDKA;1
- PMID: 19948791
- PMCID: PMC2798325
- DOI: 10.1105/tpc.109.070417
Control of cell proliferation, organ growth, and DNA damage response operate independently of dephosphorylation of the Arabidopsis Cdk1 homolog CDKA;1
Abstract
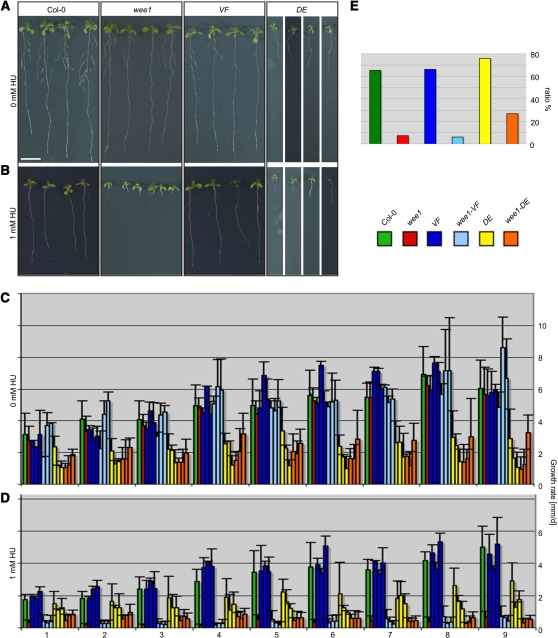
Entry into mitosis is universally controlled by cyclin-dependent kinases (CDKs). A key regulatory event in metazoans and fission yeast is CDK activation by the removal of inhibitory phosphate groups in the ATP binding pocket catalyzed by Cdc25 phosphatases. In contrast with other multicellular organisms, we show here that in the flowering plant Arabidopsis thaliana, cell cycle control does not depend on sudden changes in the phosphorylation pattern of the PSTAIRE-containing Cdk1 homolog CDKA;1. Consistently, we found that neither mutants in a previously identified CDC25 candidate gene nor plants in which it is overexpressed display cell cycle defects. Inhibitory phosphorylation of CDKs is also the key event in metazoans to arrest cell cycle progression upon DNA damage. However, we show here that the DNA damage checkpoint in Arabidopsis can also operate independently of the phosphorylation of CDKA;1. These observations reveal a surprising degree of divergence in the circuitry of highly conserved core cell cycle regulators in multicellular organisms. Based on biomathematical simulations, we propose a plant-specific model of how progression through the cell cycle could be wired in Arabidopsis.
Figures







Similar articles
-
Genetic framework of cyclin-dependent kinase function in Arabidopsis.Dev Cell. 2012 May 15;22(5):1030-40. doi: 10.1016/j.devcel.2012.02.015. Dev Cell. 2012. PMID: 22595674
-
The cyclin-dependent kinase inhibitor KRP2 controls the onset of the endoreduplication cycle during Arabidopsis leaf development through inhibition of mitotic CDKA;1 kinase complexes.Plant Cell. 2005 Jun;17(6):1723-36. doi: 10.1105/tpc.105.032383. Epub 2005 Apr 29. Plant Cell. 2005. PMID: 15863515 Free PMC article.
-
A general G1/S-phase cell-cycle control module in the flowering plant Arabidopsis thaliana.PLoS Genet. 2012;8(8):e1002847. doi: 10.1371/journal.pgen.1002847. Epub 2012 Aug 2. PLoS Genet. 2012. PMID: 22879821 Free PMC article.
-
DNA damage: Chk1 and Cdc25, more than meets the eye.Curr Opin Genet Dev. 2001 Feb;11(1):78-82. doi: 10.1016/s0959-437x(00)00160-x. Curr Opin Genet Dev. 2001. PMID: 11163155 Review.
-
[Role of CDC25 phosphatases in the control of proliferation].Pathol Biol (Paris). 2000 Apr;48(3):182-9. Pathol Biol (Paris). 2000. PMID: 10858952 Review. French.
Cited by
-
The Arabidopsis Cdk1/Cdk2 homolog CDKA;1 controls chromosome axis assembly during plant meiosis.EMBO J. 2020 Feb 3;39(3):e101625. doi: 10.15252/embj.2019101625. Epub 2019 Sep 26. EMBO J. 2020. PMID: 31556459 Free PMC article.
-
The Sos-recruitment system as a tool to analyze cellular localization of plant proteins: membrane localization of Arabidopsis thaliana PEPINO/PASTICCINO2.Mol Genet Genomics. 2010 May;283(5):439-49. doi: 10.1007/s00438-010-0528-5. Epub 2010 Mar 19. Mol Genet Genomics. 2010. PMID: 20300944
-
A distinct class of plant and animal viral proteins that disrupt mitosis by directly interrupting the mitotic entry switch Wee1-Cdc25-Cdk1.Sci Adv. 2020 May 13;6(20):eaba3418. doi: 10.1126/sciadv.aba3418. eCollection 2020 May. Sci Adv. 2020. PMID: 32426509 Free PMC article.
-
Leaf development: a cellular perspective.Front Plant Sci. 2014 Jul 31;5:362. doi: 10.3389/fpls.2014.00362. eCollection 2014. Front Plant Sci. 2014. PMID: 25132838 Free PMC article. Review.
-
The ATR-WEE1 kinase module promotes SUPPRESSOR OF GAMMA RESPONSE 1 translation to activate replication stress responses.Plant Cell. 2023 Aug 2;35(8):3021-3034. doi: 10.1093/plcell/koad126. Plant Cell. 2023. PMID: 37159556 Free PMC article.
References
-
- Amon, A., Surana, U., Muroff, I., and Nasmyth, K. (1992). Regulation of p34CDC28 tyrosine phosphorylation is not required for entry into mitosis in S. cerevisiae. Nature 355 368–371. - PubMed
-
- Araki, S., Ito, M., Soyano, T., Nishihama, R., and Machida, Y. (2004). Mitotic cyclins stimulate the activity of c-Myb-like factors for transactivation of G2/M phase-specific genes in tobacco. J. Biol. Chem. 279 32979–32988. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
