

Aquaporin 9 expression in the developing rat epididymis is modulated by steroid hormones
- PMID: 19948840
- PMCID: PMC3773695
- DOI: 10.1530/REP-09-0284
Aquaporin 9 expression in the developing rat epididymis is modulated by steroid hormones
Abstract
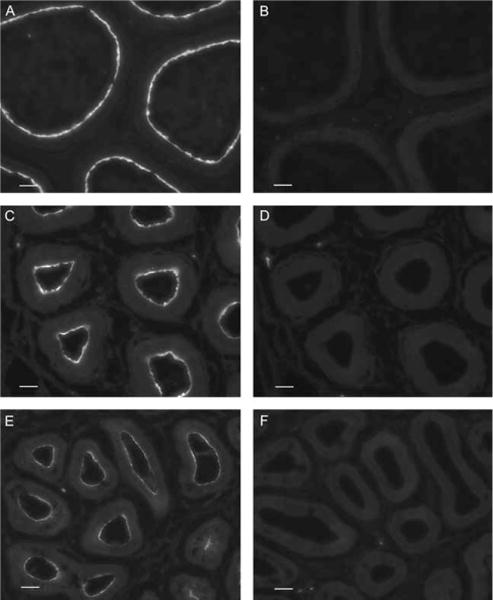
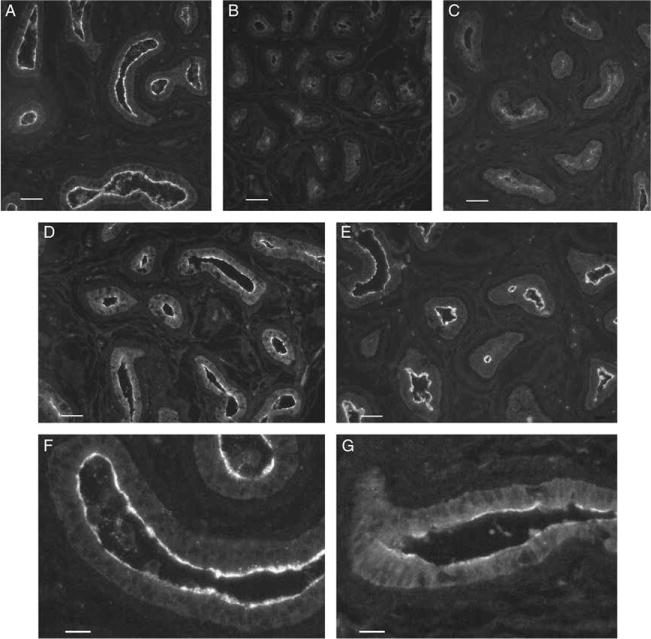
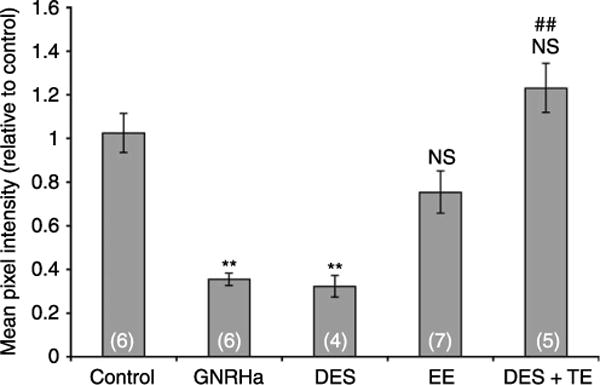
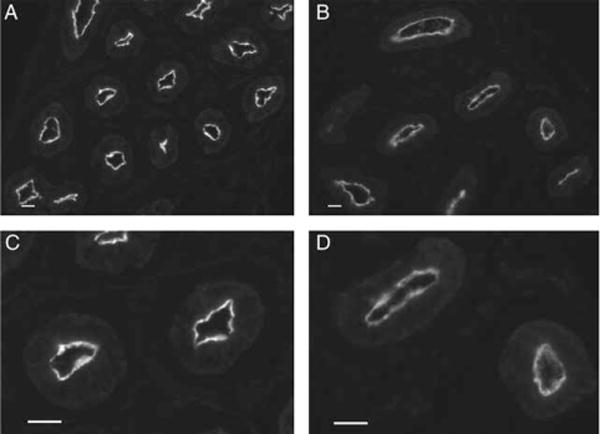
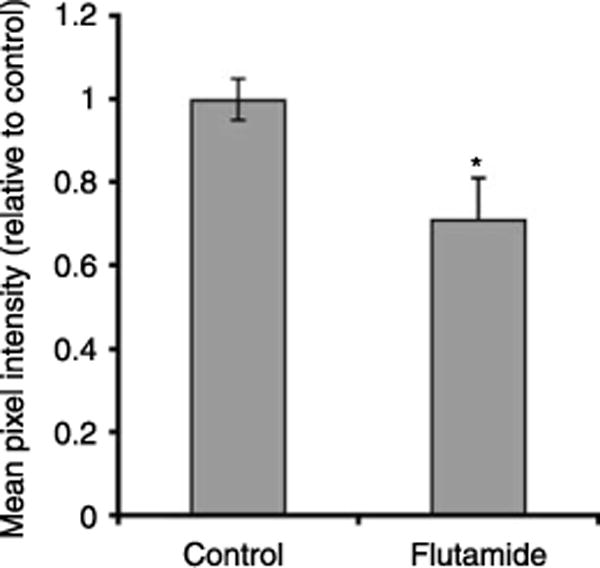
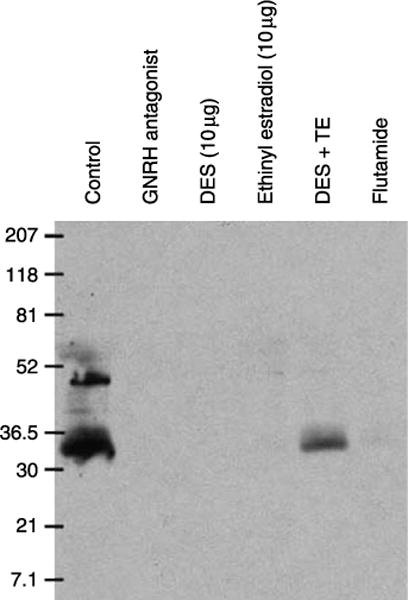
Fluid and solute transport across the epithelium of the male excurrent duct is important for sperm maturation and storage. Aquaporin 9 (AQP9), which allows permeation of water and neutral solutes, is abundant throughout the male reproductive tract, where it is expressed at the apical membrane of rat epididymal principal cells as early as at 1 week of age. We evaluated the effect of neonatal exposure to: 1) a GNRH antagonist (GNRHa); 2) diethylstilbestrol (DES); 3) ethinyl estradiol (EE); 4) DES plus testosterone (DES+TE); and 5) the anti-androgen flutamide on AQP9 expression in the epididymis of peripubertal rats. Control groups received the vehicle alone. In 25-day-old rats, quantification of the mean pixel intensity of immunofluorescence-stained sections showed a significant decrease in AQP9 staining in the apical membrane of epididymal principal cells after treatments with GNRHa, DES, or flutamide, compared to controls. These results were confirmed by western blotting. While EE induced a marked decrease in AQP9 levels by western blotting, the decrease in AQP9-associated fluorescence was not significant compared to controls. DES+TE-treated rats showed levels of AQP9 protein similar to controls, indicating maintenance of AQP9 expression by testosterone treatment in the presence of DES. Our data show that expression of AQP9 in the developing rat epididymis is downregulated by neonatal DES, GNRHa, EE, and flutamide, and that the effects mediated by estrogens can be prevented by testosterone administration.
Conflict of interest statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Figures
References
-
- Atanassova N, McKinnell C, Walker M, Turner KJ, Fisher JS, Morley M, Millar MR, Groome NP, Sharpe RM. Permanent effects of neonatal estrogen exposure in rats on reproductive hormone levels, Sertoli cell number, and the efficiency of spermatogenesis in adulthood. Endocrinology. 1999;140:5364–5373. - PubMed
-
- Atanassova N, McKinnell C, Fisher J, Sharpe RM. Neonatal treatment of rats with diethylstilboestrol (DES) induces stromal–epithelial abnormalities of the vas deferens and cauda epididymis in adulthood following delayed basal cell development. Reproduction. 2005;129:589–601. - PubMed
-
- Badran HH, Hermo LS. Expression and regulation of aquaporins 1, 8, and 9 in the testis, efferent ducts, and epididymis of adult rats and during postnatal development. Journal of Andrology. 2002;23:358–373. - PubMed
-
- Bagnis C, Marsolais M, Biemesderfer D, Laprade R, Breton S. Na+/H+-exchange activity and immunolocalization of NHE3 in rat epididymis. American Journal of Physiology Renal Physiology. 2001;280:F426–F436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
