

Contributions of Francisella tularensis subsp. novicida chitinases and Sec secretion system to biofilm formation on chitin
- PMID: 19948864
- PMCID: PMC2805214
- DOI: 10.1128/AEM.02037-09
Contributions of Francisella tularensis subsp. novicida chitinases and Sec secretion system to biofilm formation on chitin
Abstract
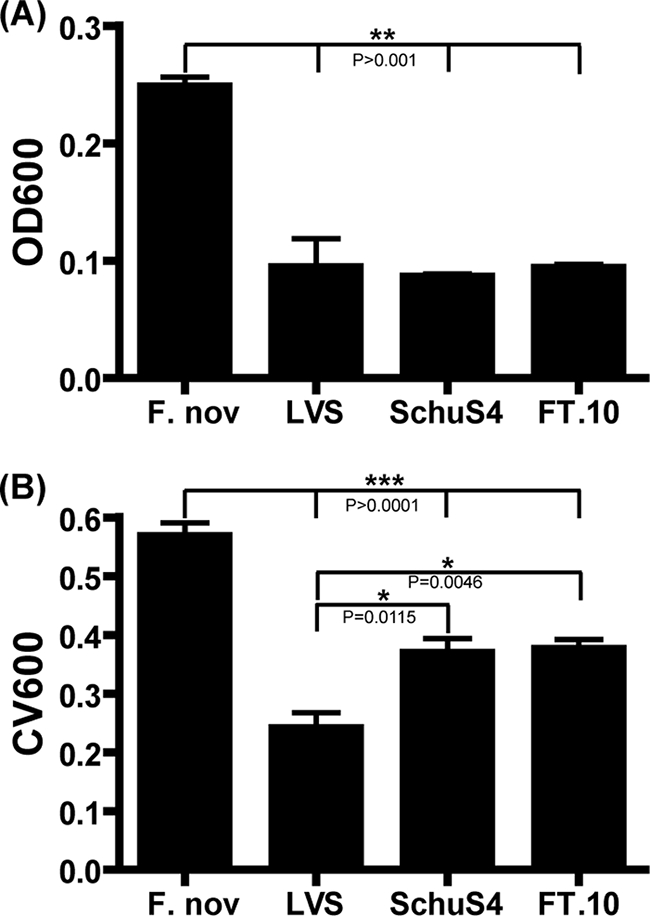
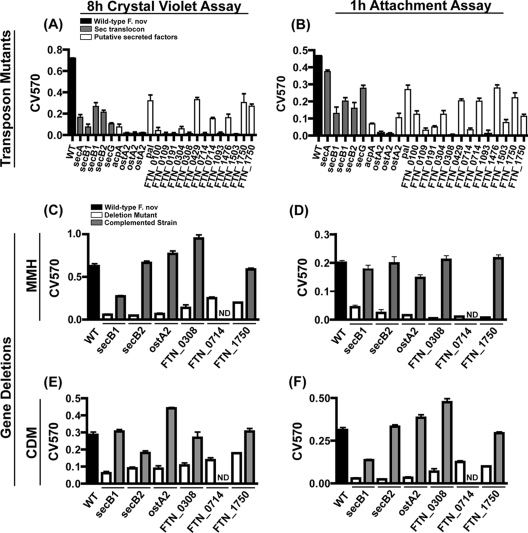
Francisella tularensis, the zoonotic cause of tularemia, can infect numerous mammals and other eukaryotes. Although studying F. tularensis pathogenesis is essential to comprehending disease, mammalian infection is just one step in the ecology of Francisella species. F. tularensis has been isolated from aquatic environments and arthropod vectors, environments in which chitin could serve as a potential carbon source and as a surface for attachment and growth. We show that F. tularensis subsp. novicida forms biofilms during the colonization of chitin surfaces. The ability of F. tularensis to persist using chitin as a sole carbon source is dependent on chitinases, since mutants lacking chiA or chiB are attenuated for chitin colonization and biofilm formation in the absence of exogenous sugar. A genetic screen for biofilm mutants identified the Sec translocon export pathway and 14 secreted proteins. We show that these genes are important for initial attachment during biofilm formation. We generated defined deletion mutants by targeting two chaperone genes (secB1 and secB2) involved in Sec-dependent secretion and four genes that encode putative secreted proteins. All of the mutants were deficient in attachment to polystyrene and chitin surfaces and for biofilm formation compared to wild-type F. novicida. In contrast, mutations in the Sec translocon and secreted factors did not affect virulence. Our data suggest that biofilm formation by F. tularensis promotes persistence on chitin surfaces. Further study of the interaction of F. tularensis with the chitin microenvironment may provide insight into the environmental survival and transmission mechanisms of this pathogen.
Figures

References
-
- Anda, P., J. Segura del Pozo, J. M. Díaz García, R. Escudero, F. J. García Peña, M. C. López Velasco, R. E. Sellek, M. R. Jiménez Chillarón, L. P. Sánchez Serrano, and J. F. Martínez Navarro. 2001. Waterborne outbreak of tularemia associated with crayfish fishing. Emerg. Infect. Dis. 7:575-582. - PMC - PubMed
-
- Balestrino, D., J. M. Ghigo, N. Charbonnel, J. A. Haagensen, and C. Forestier. 2008. The characterization of functions involved in the establishment and maturation of Klebsiella pneumoniae in vitro biofilm reveals dual roles for surface exopolysaccharides. Environ. Microbiol. 10:685-701. - PubMed
-
- Bartnicki-Garcia, S. 1968. Cell wall chemistry, morphogenesis, and taxonomy of fungi. Annu. Rev. Microbiol. 22:87-108. - PubMed
-
- Bassler, B. L., C. Yu, Y. C. Lee, and S. Roseman. 1991. Chitin utilization by marine bacteria. Degradation and catabolism of chitin oligosaccharides by Vibrio furnissii. J. Biol. Chem. 266:24276-24286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
