

FRQ-interacting RNA helicase mediates negative and positive feedback in the Neurospora circadian clock
- PMID: 19948888
- PMCID: PMC2828717
- DOI: 10.1534/genetics.109.111393
FRQ-interacting RNA helicase mediates negative and positive feedback in the Neurospora circadian clock
Abstract
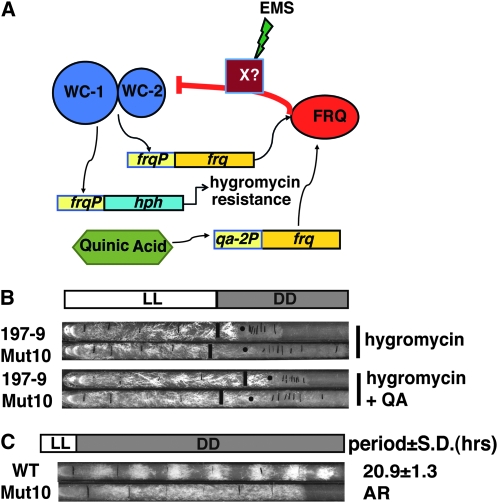
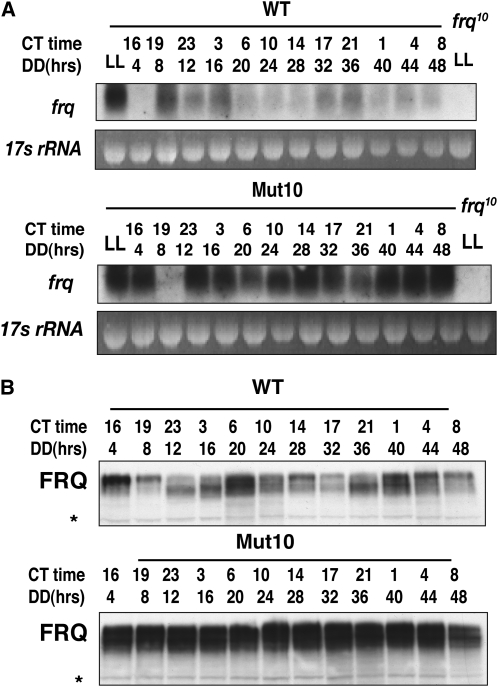
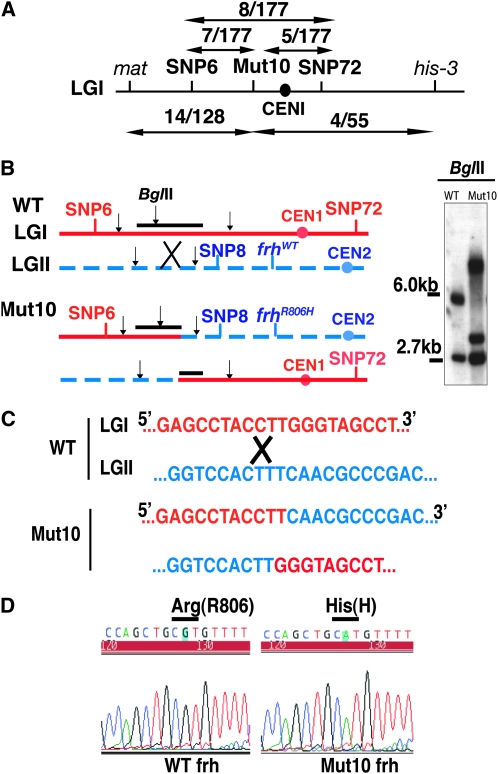
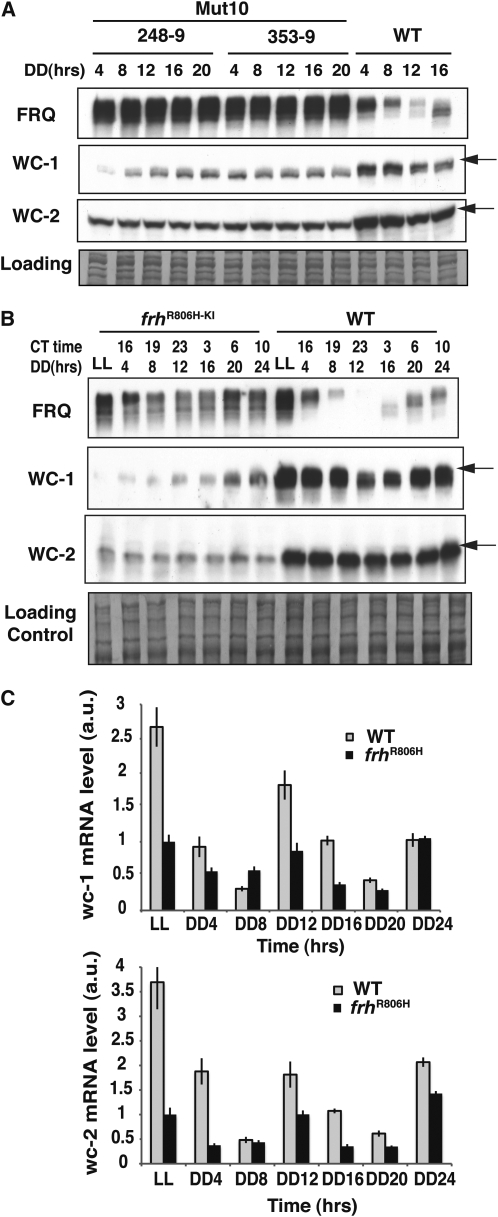
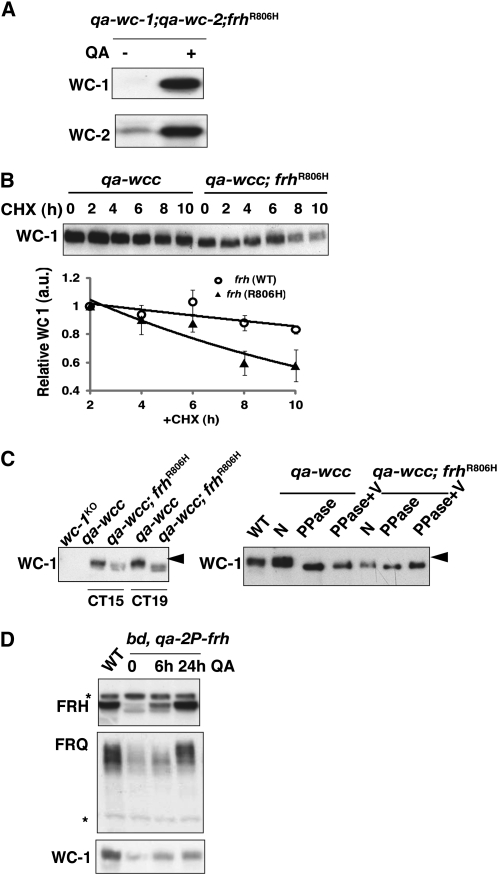
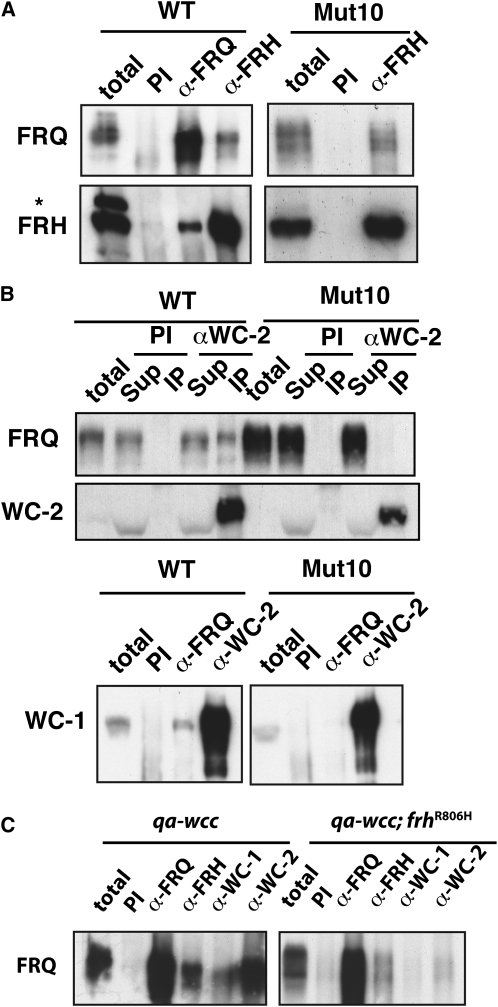
The Neurospora circadian oscillator comprises FREQUENCY (FRQ) and its transcription activator, the White Collar Complex (WCC). Repression of WCC's transcriptional activity by FRQ via negative feedback is indispensable for clock function. An unbiased genetic screen that targeted mutants with defects in negative feedback regulation yielded a fully viable arrhythmic strain bearing a novel allele of FRQ-interacting RNA helicase (frh), an essential gene that encodes a putative exosome component protein. In the allele, frh(R806H), clock function is completely disturbed, while roles of FRQ-interacting RNA helicase (FRH) essential for viability are left intact. FRH(R806H) still interacts with FRQ, but interaction between the FRQ-FRH(R806H) complex (FFC) and WCC is severely affected. Phosphorylation of WC-1 is reduced in the mutant leading to constantly elevated WCC activity, which breaks the negative feedback loop. WCC levels are considerably reduced in the mutant, especially those of WC-1, consistent both with loss of positive feedback (FRQ-dependent WC-1 stabilization) and with a reduced level of the FRQ-mediated WCC phosphorylation that leads to high WCC activity accompanied by rapid transcription-associated turnover. FRH overexpression promotes WC-1 accumulation, confirming that FRH together with FRQ plays a role in WC-1 stabilization. Identification of a viable allele of frh, displaying virtually complete loss of both negative and positive circadian feedback, positions FRH as a core component of the central oscillator that is permissive for rhythmicity but appears not to modulate periodicity. Moreover, the results suggest that there are clock-specific roles for FRH that are distinct from the predicted essential exosome-associated functions for the protein.
Figures

References
-
- Aronson, B. D., K. A. Johnson, J. J. Loros and J. C. Dunlap, 1994. Negative feedback defining a circadian clock: autoregulation of the clock gene frequency. Science 263 1578–1584. - PubMed
-
- Brunner, M., and K. Kaldi, 2008. Interlocked feedback loops of the circadian clock of Neurospora crassa. Mol. Microbiol. 68 255–262. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
