

Structural insights into tail-anchored protein binding and membrane insertion by Get3
- PMID: 19948960
- PMCID: PMC2795547
- DOI: 10.1073/pnas.0910223106
Structural insights into tail-anchored protein binding and membrane insertion by Get3
Abstract
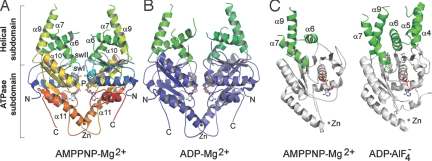
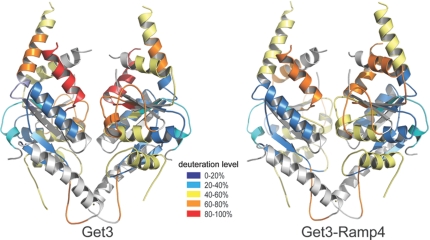
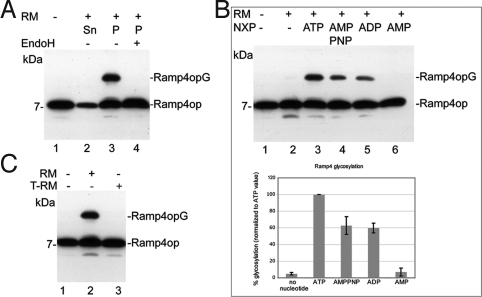
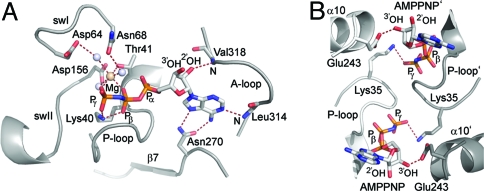
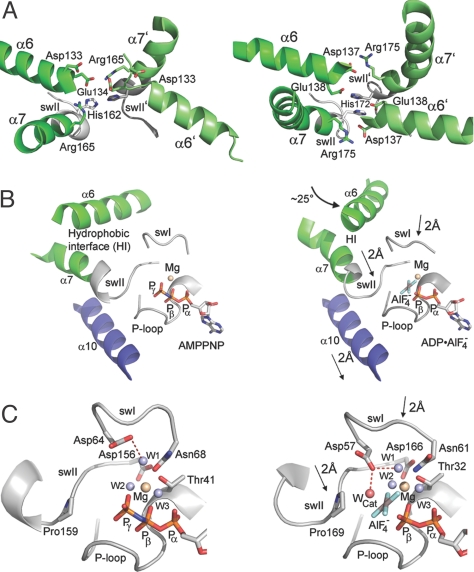
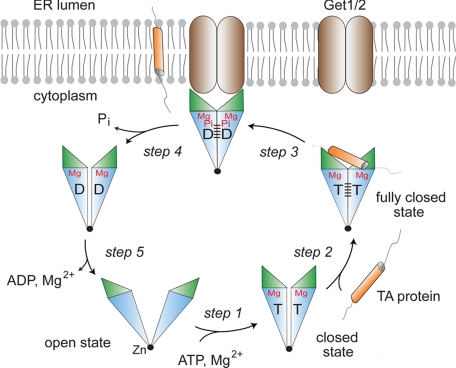
Tail-anchored (TA) membrane proteins are involved in a variety of important cellular functions, including membrane fusion, protein translocation, and apoptosis. The ATPase Get3 (Asna1, TRC40) was identified recently as the endoplasmic reticulum targeting factor of TA proteins. Get3 consists of an ATPase and alpha-helical subdomain enriched in methionine and glycine residues. We present structural and biochemical analyses of Get3 alone as well as in complex with a TA protein, ribosome-associated membrane protein 4 (Ramp4). The ATPase domains form an extensive dimer interface that encloses 2 nucleotides in a head-to-head orientation and a zinc ion. Amide proton exchange mass spectrometry shows that the alpha-helical subdomain of Get3 displays considerable flexibility in solution and maps the TA protein-binding site to the alpha-helical subdomain. The non-hydrolyzable ATP analogue AMPPNP-Mg(2+)- and ADP-Mg(2+)-bound crystal structures representing the pre- and posthydrolysis states are both in a closed form. In the absence of a TA protein cargo, ATP hydrolysis does not seem to be possible. Comparison with the ADP.AlF(4)(-)-bound structure representing the transition state (Mateja A, et al. (2009) Nature 461:361-366) indicates how the presence of a TA protein is communicated to the ATP-binding site. In vitro membrane insertion studies show that recombinant Get3 inserts Ramp4 in a nucleotide- and receptor-dependent manner. Although ATP hydrolysis is not required for Ramp4 insertion per se, it seems to be required for efficient insertion. We postulate that ATP hydrolysis is needed to release Get3 from its receptor. Taken together, our results provide mechanistic insights into posttranslational targeting of TA membrane proteins by Get3.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Get1 stabilizes an open dimer conformation of get3 ATPase by binding two distinct interfaces.J Mol Biol. 2012 Sep 21;422(3):366-75. doi: 10.1016/j.jmb.2012.05.045. Epub 2012 Jun 7. J Mol Biol. 2012. PMID: 22684149
-
Structural insight into the membrane insertion of tail-anchored proteins by Get3.Genes Cells. 2010 Jan;15(1):29-41. doi: 10.1111/j.1365-2443.2009.01362.x. Epub 2009 Dec 15. Genes Cells. 2010. PMID: 20015340
-
The structural basis of tail-anchored membrane protein recognition by Get3.Nature. 2009 Sep 17;461(7262):361-6. doi: 10.1038/nature08319. Epub 2009 Aug 12. Nature. 2009. PMID: 19675567 Free PMC article.
-
The natural history of Get3-like chaperones.Traffic. 2019 May;20(5):311-324. doi: 10.1111/tra.12643. Traffic. 2019. PMID: 30972921 Free PMC article. Review.
-
Structures of Get3, Get4, and Get5 provide new models for TA membrane protein targeting.Structure. 2010 Aug 11;18(8):897-902. doi: 10.1016/j.str.2010.07.003. Structure. 2010. PMID: 20696390 Free PMC article. Review.
Cited by
-
Nuclear import of UBL-domain protein Mdy2 is required for heat-induced stress response in Saccharomyces cerevisiae.PLoS One. 2012;7(12):e52956. doi: 10.1371/journal.pone.0052956. Epub 2012 Dec 28. PLoS One. 2012. PMID: 23285234 Free PMC article.
-
Pathways of arsenic uptake and efflux.Curr Top Membr. 2012;69:325-58. doi: 10.1016/B978-0-12-394390-3.00012-4. Curr Top Membr. 2012. PMID: 23046656 Free PMC article.
-
SIMIBI twins in protein targeting and localization.Nat Struct Mol Biol. 2013 Jul;20(7):776-80. doi: 10.1038/nsmb.2605. Nat Struct Mol Biol. 2013. PMID: 23823071
-
Consistent mutational paths predict eukaryotic thermostability.BMC Evol Biol. 2013 Jan 10;13:7. doi: 10.1186/1471-2148-13-7. BMC Evol Biol. 2013. PMID: 23305080 Free PMC article.
-
Guiding tail-anchored membrane proteins to the endoplasmic reticulum in a chaperone cascade.J Biol Chem. 2019 Nov 8;294(45):16577-16586. doi: 10.1074/jbc.REV119.006197. Epub 2019 Oct 1. J Biol Chem. 2019. PMID: 31575659 Free PMC article. Review.
References
-
- Rapoport TA, Jungnickel B, Kutay U. Protein transport across the eukaryotic endoplasmic reticulum and bacterial inner membranes. Annu Rev Biochem. 1996;65:271–303. - PubMed
-
- Cross BC, Sinning I, Luirink J, High S. Delivering proteins for export from the cytosol. Nat Rev Mol Cell Biol. 2009;10:255–264. - PubMed
-
- Keenan RJ, Freymann DM, Stroud RM, Walter P. The signal recognition particle. Annu Rev Biochem. 2001;70:755–775. - PubMed
-
- Wild K, Rosendal KR, Sinning I. A structural step into the SRP cycle. Mol Microbiol. 2004;53:357–363. - PubMed
-
- Rapoport TA. Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes. Nature. 2007;450:663–669. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
