

blaCMY-2-positive IncA/C plasmids from Escherichia coli and Salmonella enterica are a distinct component of a larger lineage of plasmids
- PMID: 19949054
- PMCID: PMC2812137
- DOI: 10.1128/AAC.00055-09
blaCMY-2-positive IncA/C plasmids from Escherichia coli and Salmonella enterica are a distinct component of a larger lineage of plasmids
Abstract
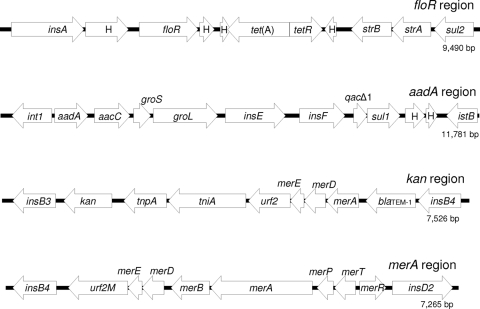
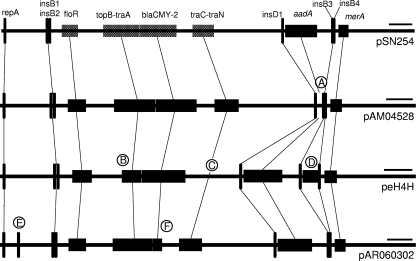
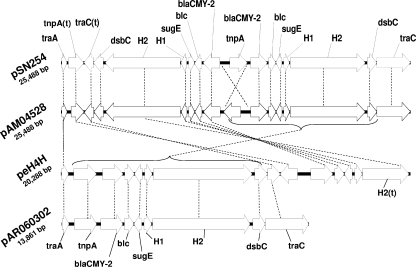
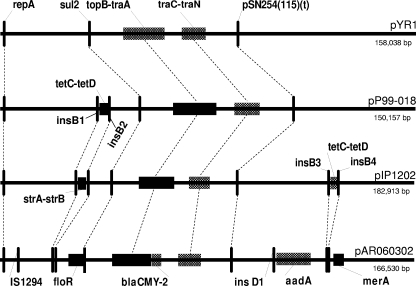
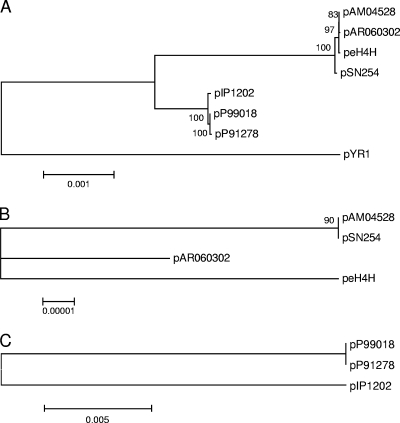
Large multidrug resistance plasmids of the A/C incompatibility complex (IncA/C) have been found in a diverse group of Gram-negative commensal and pathogenic bacteria. We present three completed sequences from IncA/C plasmids that originated from Escherichia coli (cattle) and Salmonella enterica serovar Newport (human) and that carry the cephamycinase gene blaCMY-2. These large plasmids (148 to 166 kbp) share extensive sequence identity and synteny. The most divergent plasmid, peH4H, has lost several conjugation-related genes and has gained a kanamycin resistance region. Two of the plasmids (pAM04528 and peH4H) harbor two copies of blaCMY-2, while the third plasmid (pAR060302) harbors a single copy of the gene. The majority of single-nucleotide polymorphisms comprise nonsynonymous mutations in floR. A comparative analysis of these plasmids with five other published IncA/C plasmids showed that the blaCMY-2 plasmids from E. coli and S. enterica are genetically distinct from those originating from Yersinia pestis and Photobacterium damselae and distal to one originating from Yersinia ruckeri. While the overall similarity of these plasmids supports the likelihood of recent movements among E. coli and S. enterica hosts, their greater divergence from Y. pestis or Y. ruckeri suggests less recent plasmid transfer among these pathogen groups.
Figures
Comment in
-
ISCR elements are key players in IncA/C plasmid evolution.Antimicrob Agents Chemother. 2010 Aug;54(8):3534; author reply 3534. doi: 10.1128/AAC.00383-10. Antimicrob Agents Chemother. 2010. PMID: 20634542 Free PMC article. No abstract available.
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Bauer, A. W., W. M. Kirby, J. C. Sherris, and M. Turck. 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 45:493-496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
