

Implications of functional anatomy on information processing in the deep cerebellar nuclei
- PMID: 19949453
- PMCID: PMC2783015
- DOI: 10.3389/neuro.03.014.2009
Implications of functional anatomy on information processing in the deep cerebellar nuclei
Abstract
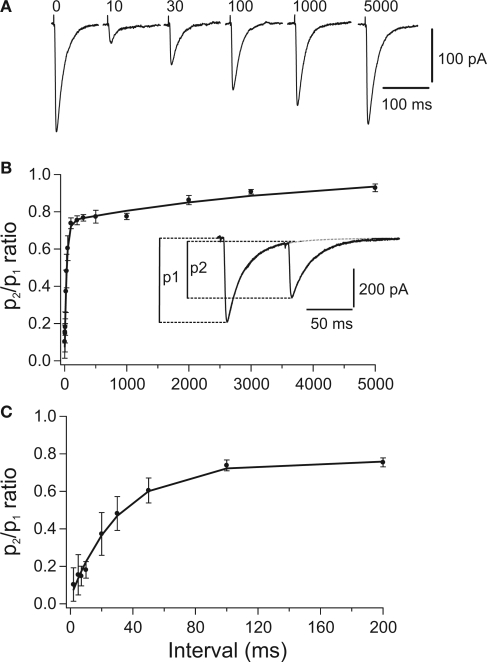
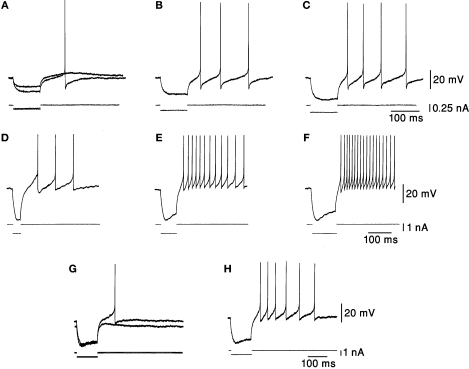
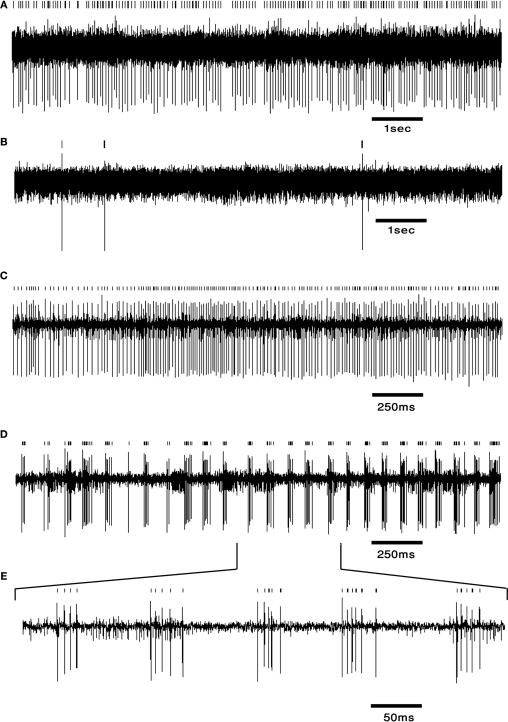
The cerebellum has been implicated as a major player in producing temporal acuity. Theories of cerebellar timing typically emphasize the role of the cerebellar cortex while overlooking the role of the deep cerebellar nuclei (DCN) that provide the sole output of the cerebellum. Here we review anatomical and electrophysiological studies to shed light on the DCN's ability to support temporal pattern generation in the cerebellum. Specifically, we examine data on the structure of the DCN, the biophysical properties of DCN neurons and properties of the afferent systems to evaluate their contribution to DCN firing patterns. In addition, we manipulate one of the afferent structures, the inferior olive (IO), using systemic harmaline injection to test for a network effect on activity of single DCN neurons in freely moving animals. Harmaline induces a rhythmic firing pattern of short bursts on a quiescent background at about 8 Hz. Other neurons become quiescent for long periods (seconds to minutes). The observed patterns indicate that the major effect harmaline exerts on the DCN is carried indirectly by the inhibitory Purkinje cells (PCs) activated by the IO, rather than by direct olivary excitation. Moreover, we suggest that the DCN response profile is determined primarily by the number of concurrently active PCs, their firing rate and the level of synchrony occurring in their transitions between continuous firing and quiescence. We argue that DCN neurons faithfully transfer temporal patterns resulting from strong correlations in PCs state transitions, while largely ignoring the timing of simple spikes from individual PCs. Future research should aim at quantifying the contribution of PC state transitions to DCN activity, and the interplay between the different afferent systems that drive DCN activity.
Keywords: cerebellar nuclei; chronic recording; harmaline; inferior olive; rebound firing; short-term depression; temporal patterns.
Figures
References
-
- Aizenman C. D., Linden D. J. (1999). Regulation of the rebound depolarization and spontaneous firing patterns of deep nuclear neurons in slices of rat cerebellum. J. Neurophysiol. 82, 1697–1709 - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous