

Nuclear respiratory factor 2 induces the expression of many but not all human proteins acting in mitochondrial DNA transcription and replication
- PMID: 19951946
- PMCID: PMC2823536
- DOI: 10.1074/jbc.M109.044305
Nuclear respiratory factor 2 induces the expression of many but not all human proteins acting in mitochondrial DNA transcription and replication
Abstract
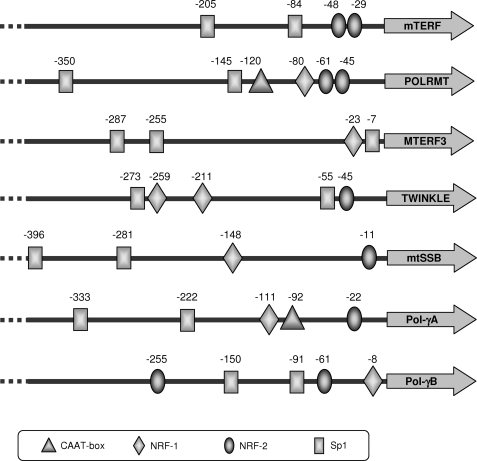
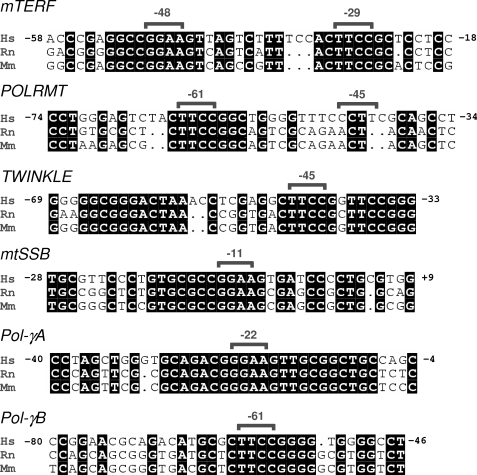
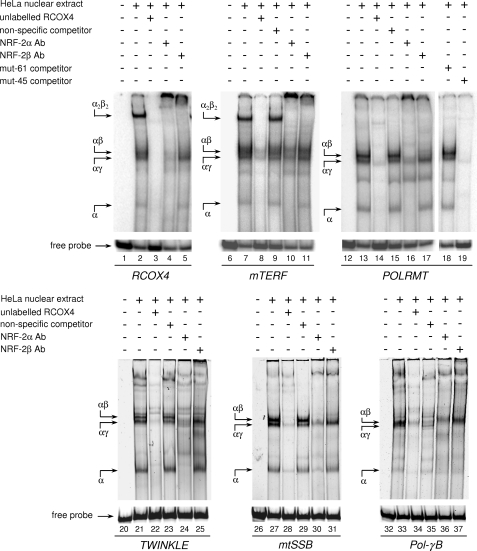
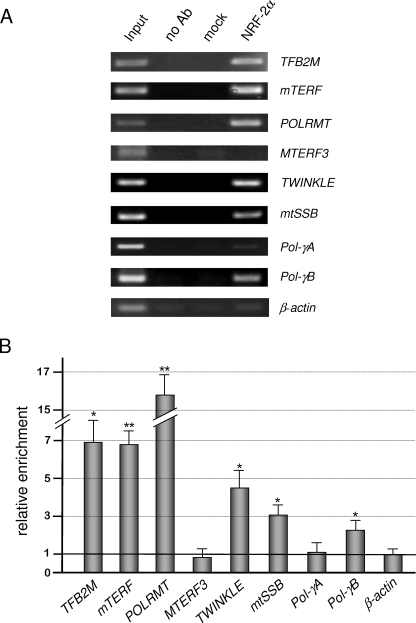
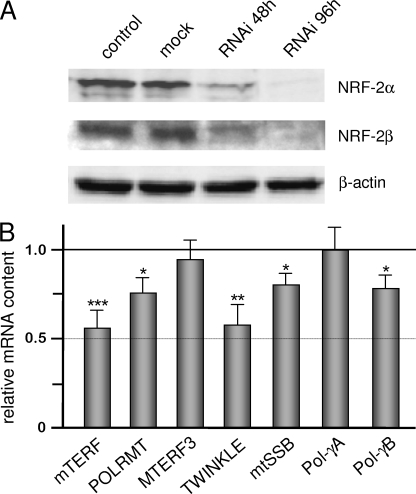
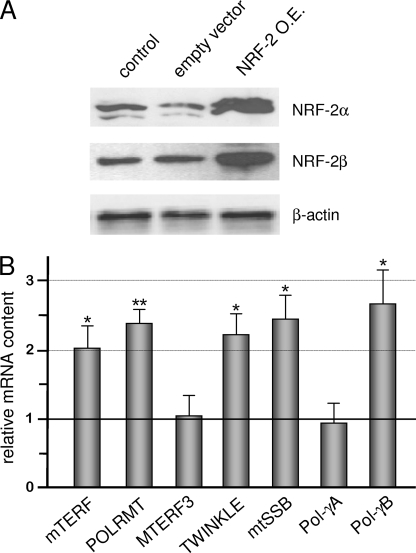
In mammals, NRF-2 (nuclear respiratory factor 2), also named GA-binding protein, is an Ets family transcription factor that controls many genes involved in cell cycle progression and protein synthesis as well as in mitochondrial biogenesis. In this paper, we analyzed the role of NRF-2 in the regulation of human genes involved in mitochondrial DNA transcription and replication. By a combination of bioinformatic and biochemical approaches, we found that the factor binds in vitro and in vivo to the proximal promoter region of the genes coding for the transcription termination factor mTERF, the RNA polymerase POLRMT, the B subunit of the DNA polymerase-gamma, the DNA helicase TWINKLE, and the single-stranded DNA-binding protein mtSSB. The role of NRF-2 in modulating the expression of those genes was further established by RNA interference and overexpression strategies. On the contrary, we found that NRF-2 does not control the genes for the subunit A of DNA polymerase-gamma and for the transcription repressor MTERF3; we suggest that these genes are under regulatory mechanisms that do not involve NRF proteins. Since NRFs are known to positively control the expression of transcription-activating proteins, the novelty emerging from our data is that proteins playing antithetical roles in mitochondrial DNA transcription, namely activators and repressors, are under different regulatory pathways. Finally, we developed a more stringent consensus with respect to the general consensus of NRF-2/GA-binding protein when searching for NRF-2 binding sites in the promoter of mitochondrial proteins.
Figures

Similar articles
-
Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: a potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis.Proc Natl Acad Sci U S A. 1994 Feb 15;91(4):1309-13. doi: 10.1073/pnas.91.4.1309. Proc Natl Acad Sci U S A. 1994. PMID: 8108407 Free PMC article.
-
In vitro methylation of nuclear respiratory factor-2 binding sites suppresses the promoter activity of the human TOMM70 gene.Gene. 2008 Dec 31;427(1-2):58-64. doi: 10.1016/j.gene.2008.09.016. Epub 2008 Sep 25. Gene. 2008. PMID: 18852034
-
Estradiol stimulates transcription of nuclear respiratory factor-1 and increases mitochondrial biogenesis.Mol Endocrinol. 2008 Mar;22(3):609-22. doi: 10.1210/me.2007-0029. Epub 2007 Nov 29. Mol Endocrinol. 2008. PMID: 18048642 Free PMC article.
-
The interface of transcription and DNA replication in the mitochondria.Biochim Biophys Acta. 2012 Sep-Oct;1819(9-10):970-8. doi: 10.1016/j.bbagrm.2011.12.005. Epub 2011 Dec 20. Biochim Biophys Acta. 2012. PMID: 22207204 Free PMC article. Review.
-
Mitochondrial transcription and its regulation in mammalian cells.Trends Biochem Sci. 2007 Mar;32(3):111-7. doi: 10.1016/j.tibs.2007.01.003. Epub 2007 Feb 8. Trends Biochem Sci. 2007. PMID: 17291767 Review.
Cited by
-
eIF2α phosphorylation-ATF4 axis-mediated transcriptional reprogramming mitigates mitochondrial impairment during ER stress.Mol Cells. 2025 Feb;48(2):100176. doi: 10.1016/j.mocell.2024.100176. Epub 2025 Jan 3. Mol Cells. 2025. PMID: 39756584 Free PMC article.
-
Harnessing mitochondrial biogenesis to combat acute kidney injury: Current insights and futuredirections.Genes Dis. 2025 Apr 15;12(6):101645. doi: 10.1016/j.gendis.2025.101645. eCollection 2025 Nov. Genes Dis. 2025. PMID: 40821125 Free PMC article. Review.
-
Opa1 is required for proper mitochondrial metabolism in early development.PLoS One. 2013;8(3):e59218. doi: 10.1371/journal.pone.0059218. Epub 2013 Mar 14. PLoS One. 2013. PMID: 23516612 Free PMC article.
-
Mitochondrial network remodeling: an important feature of myogenesis and skeletal muscle regeneration.Cell Mol Life Sci. 2021 May;78(10):4653-4675. doi: 10.1007/s00018-021-03807-9. Epub 2021 Mar 22. Cell Mol Life Sci. 2021. PMID: 33751143 Free PMC article. Review.
-
Structure, mechanism, and regulation of mitochondrial DNA transcription initiation.J Biol Chem. 2020 Dec 25;295(52):18406-18425. doi: 10.1074/jbc.REV120.011202. Epub 2020 Oct 30. J Biol Chem. 2020. PMID: 33127643 Free PMC article. Review.
References
-
- Falkenberg M., Larsson N. G., Gustafsson C. M. (2007) Annu. Rev. Biochem. 76, 679–699 - PubMed
-
- Martin M., Cho J., Cesare A. J., Griffith J. D., Attardi G. (2005) Cell 123, 1227–1240 - PubMed
-
- Park C. B., Asin-Cayuela J., Cámara Y., Shi Y., Pellegrini M., Gaspari M., Wibom R., Hultenby K., Erdjument-Bromage H., Tempst P., Falkenberg M., Gustafsson C. M., Larsson N. G. (2007) Cell 130, 273–285 - PubMed
-
- Roberti M., Polosa P. L., Bruni F., Manzari C., Deceglie S., Gadaleta M. N., Cantatore P. (2009) Biochim. Biophys. Acta 1787, 303–311 - PubMed
-
- Graziewicz M. A., Longley M. J., Copeland W. C. (2006) Chem. Rev. 106, 383–405 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
