

Review
doi: 10.1101/gad.1850809.
Dividing cellular asymmetry: asymmetric cell division and its implications for stem cells and cancer
Affiliations
- PMID: 19952104
- PMCID: PMC2788323
- DOI: 10.1101/gad.1850809
Item in Clipboard
Review
Dividing cellular asymmetry: asymmetric cell division and its implications for stem cells and cancer
Genes Dev.
.
Abstract
Cell division is commonly thought to involve the equal distribution of cellular components into the two daughter cells. During many cell divisions, however, proteins, membrane compartments, organelles, or even DNA are asymmetrically distributed between the two daughter cells. Here, we review the various types of asymmetries that have been described in yeast and in animal cells. Asymmetric segregation of protein determinants is particularly relevant for stem cell biology. We summarize the relevance of asymmetric cell divisions in various stem cell systems and discuss why defects in asymmetric cell division can lead to the formation of tumors.
Figures
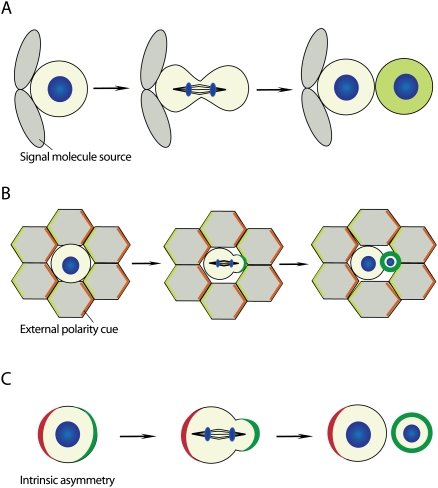
Modes of asymmetric cell division. (A) Asymmetric cell fate specification is regulated by a niche-derived signal. Cells that contact the niche retain their identity, whereas cells that become detached from the niche after division adopt a different cell fate. (B) External polarity induces the asymmetric localization of cell fate determinants (green). (C) Intrinsic asymmetry localizes polarity proteins (red), which instruct cell fate determinants (green) to segregate asymmetrically during mitosis in the absence of extracellular cues (DNA, blue).
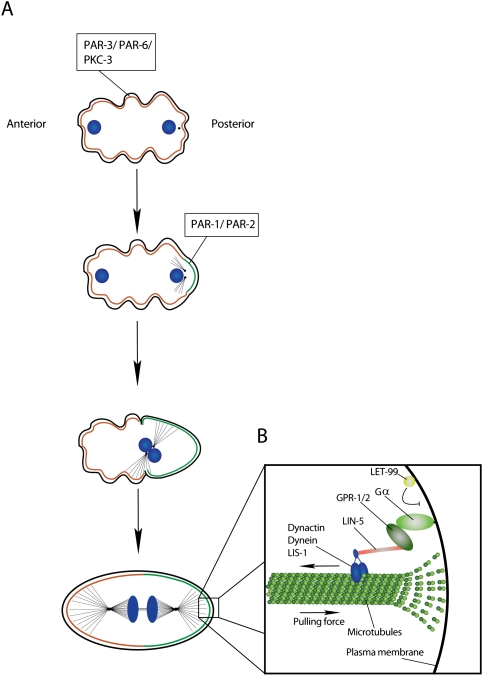
Asymmetric division of the C. elegans one-cell embryo. (A) Separation of the PAR-3/PAR-6/PKC-3 and PAR-1/PAR-2 occupied cortical domains. (B) The mitotic spindle is anchored along the anterior–posterior axis. LIN-5 and GPR1-/2 link the spindle via simultaneous binding to the microtubule-associated Dynactin/Dynein/LIS-1 complex and membrane-anchored Gα (DNA, blue).
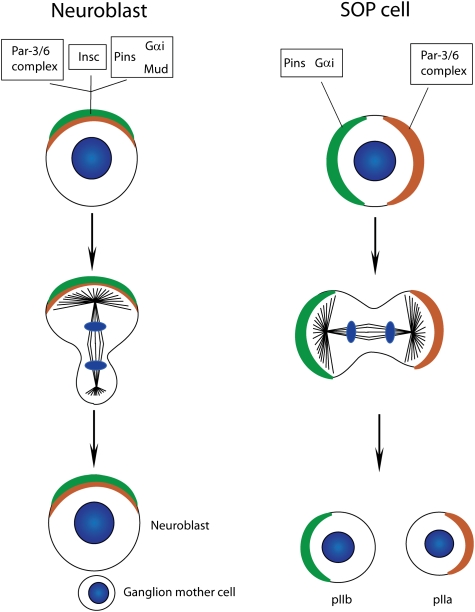
Differences in protein localization between Drosophila neuroblasts and SOP cells. The Par-6/Par-3 and the Pins/Gαi/Mud complex colocalize at the apical cortex of Drosophila neuroblasts. Pins/Gαi/Mud are recruited to the apical cortex through the presence of Insc, which simultaneously binds members of both complexes. These complexes localize to opposite cortical domains in SOP cells, as the linker molecule Insc is not expressed in this cell type (DNA, blue).
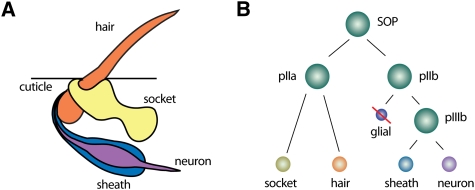
The Drosophila ES organ as a model system for asymmetric cell division. (A) The Drosophila ES organs consist of two outer (hair and socket) and two inner (neuron and sheath) cells. (B) SOP cells divide asymmetrically in a stem cell-like fashion to generate the various cells of the ES organ. Note that the glial cell undergoes programmed cell death.
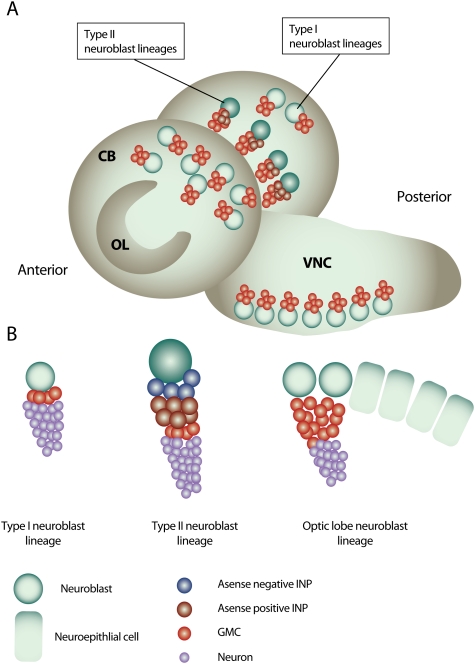
Neurogenesis in the Drosophila larval brain. (A) The Drosophila third instar larval brain contains three main neurogenic regions: the OL neurogenic center, located at the lateral surface of the two brain hemispheres, and the CB neurogenic center, which is located medially of the OL and descends to the VNC on the anterior side of the brain. (B) Type I neuroblast lineages constitute the majority of neuroblast lineages in the CB and VNC. A type I neuroblast gives rise to another neuroblast and a GMC that terminally divides to produce two neurons. Type II neuroblasts are situated on the medial, posterior surface of the brain lobes and give rise to transit-amplifying INP cells. The neuroblasts of the OL originate from OL neuroepithelial cells.
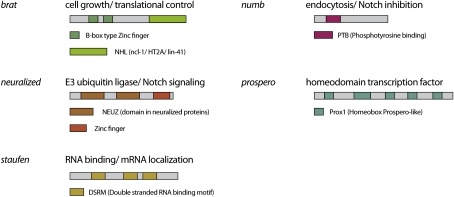
Domain architecture and main function of Drosophila cell fate determinants. Asymmetric localization and segregation of these proteins upon cell division requires the action of Par proteins.
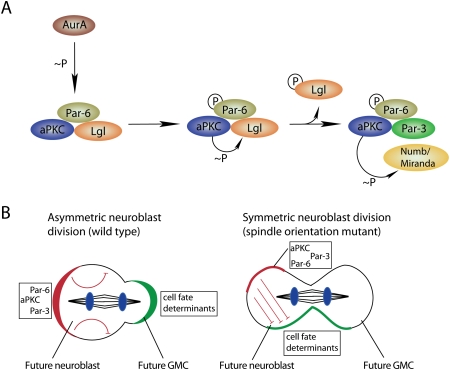
The apical domain regulates cell fate determinant localization and activity. (A) Aurora-A (AurA)-induced complex remodeling leads to the phosphorylation-mediated exclusion of the Lgl form and entry of Par-3 into the Par complex. This complex remodelling is associated with an alteration of the aPKC substrate specificity toward Numb and Miranda, which are hence excluded from the cortical domain occupied by aPKC. (B) The ratio of apical/basal determinants specifies cellular identity. Asymmetric neuroblast divisions can occur, although both daughter cells inherit cell fate determinants (green) that are inhibited by aPKC (red) in one daughter cell (DNA, blue).
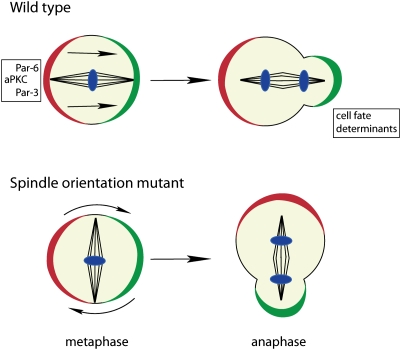
Spindle orientation defects in metaphase are frequently corrected by the “telophase rescue” pathway in anaphase/telophase. This rescue pathway restores the correct cell fate determinant segregation in late cell cycle phases in a majority of divisions in spindle orientation mutants (DNA, blue).
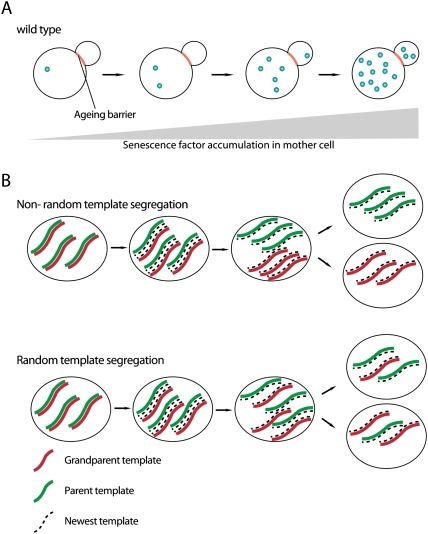
Mitotic asymmetries of senescence factors and DNA. (A) Aging/senescence factors are selectively retained in the mother cell and accumulate during the replicative life span, leading to cellular senescence and death of the mother. Daughter cells born later inherit small amounts of senescence factors, leading to a shortened replicative life span compared with younger daughter cells. (B) Asymmetric versus symmetric segregation of DNA template strands during cell division.
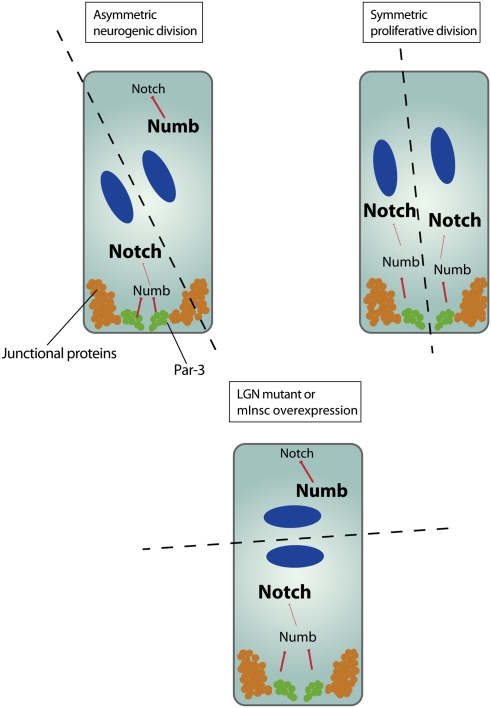
Model for cell fate specification in the vertebrate neural stem cells. Neural progenitor cells in the developing brain undergo either neurogenic divisions (in which another progenitor cell and a differentiating neuron are produced) or proliferative divisions (which result in the formation of two progenitor cells). Par-3 is partitioned unequally during neurogenic divisions through cleavage plane orientation (dashed line), and induces differential Notch signaling in the daughter cells by inhibiting Numb, resulting in high levels of Notch signaling in the future progenitor cell and low levels in the future neuron (DNA, blue).
References
-
- Abrash EB, Bergmann DC. Asymmetric cell divisions: A view from plant development. Dev Cell. 2009;16:783–796. - PubMed
-
- Afshar K, Willard FS, Colombo K, Johnston CA, McCudden CR, Siderovski DP, Gonczy P. RIC-8 is required for GPR-1/2-dependent Gα function during asymmetric division of C. elegans embryos. Cell. 2004;119:219–230. - PubMed
-
- Afshar K, Willard FS, Colombo K, Siderovski DP, Gonczy P. Cortical localization of the Gα protein GPA-16 requires RIC-8 function during C. elegans asymmetric cell division. Development. 2005;132:4449–4459. - PubMed
-
- Aguilaniu H, Gustafsson L, Rigoulet M, Nystrom T. Protein oxidation in G0 cells of Saccharomyces cerevisiae depends on the state rather than rate of respiration and is enhanced in pos9 but not yap1 mutants. J Biol Chem. 2001;276:35396–35404. - PubMed
-
- Aguilaniu H, Gustafsson L, Rigoulet M, Nystrom T. Asymmetric inheritance of oxidatively damaged proteins during cytokinesis. Science. 2003;299:1751–1753. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical