

YAP1 is amplified and up-regulated in hedgehog-associated medulloblastomas and mediates Sonic hedgehog-driven neural precursor proliferation
- PMID: 19952108
- PMCID: PMC2788333
- DOI: 10.1101/gad.1824509
YAP1 is amplified and up-regulated in hedgehog-associated medulloblastomas and mediates Sonic hedgehog-driven neural precursor proliferation
Abstract
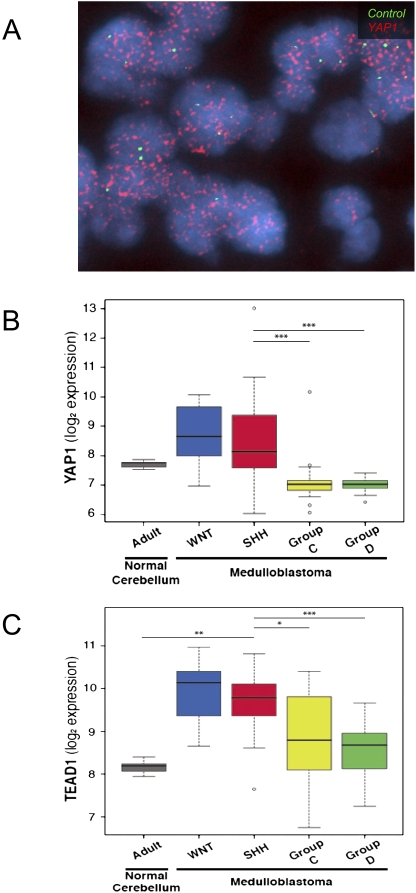
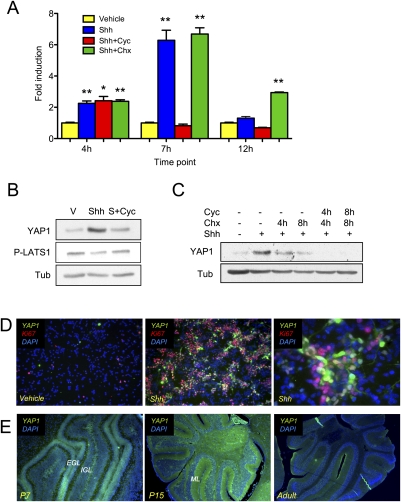
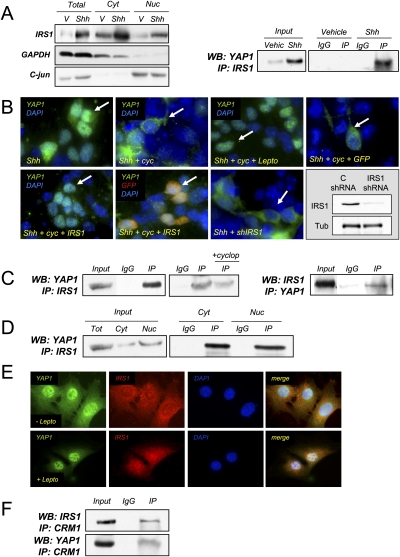
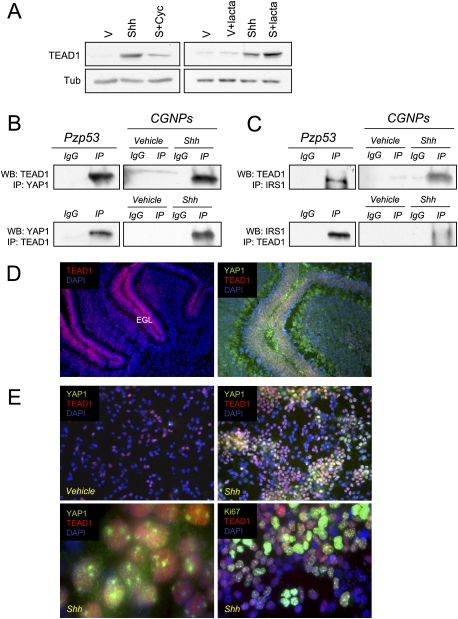
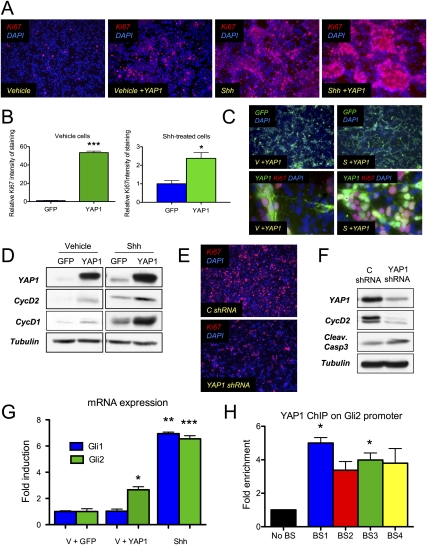
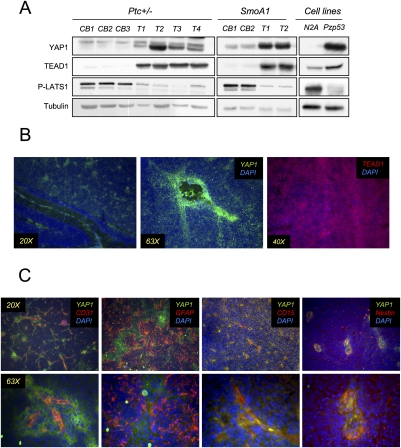
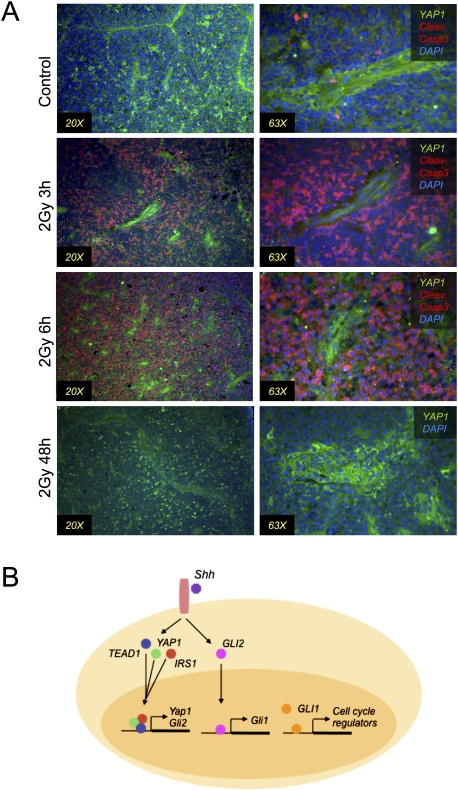
Medulloblastoma is the most common solid malignancy of childhood, with treatment side effects reducing survivors' quality of life and lethality being associated with tumor recurrence. Activation of the Sonic hedgehog (Shh) signaling pathway is implicated in human medulloblastomas. Cerebellar granule neuron precursors (CGNPs) depend on signaling by the morphogen Shh for expansion during development, and have been suggested as a cell of origin for certain medulloblastomas. Mechanisms contributing to Shh pathway-mediated proliferation and transformation remain poorly understood. We investigated interactions between Shh signaling and the recently described tumor-suppressive Hippo pathway in the developing brain and medulloblastomas. We report up-regulation of the oncogenic transcriptional coactivator yes-associated protein 1 (YAP1), which is negatively regulated by the Hippo pathway, in human medulloblastomas with aberrant Shh signaling. Consistent with conserved mechanisms between brain tumorigenesis and development, Shh induces YAP1 expression in CGNPs. Shh also promotes YAP1 nuclear localization in CGNPs, and YAP1 can drive CGNP proliferation. Furthermore, YAP1 is found in cells of the perivascular niche, where proposed tumor-repopulating cells reside. Post-irradiation, YAP1 was found in newly growing tumor cells. These findings implicate YAP1 as a new Shh effector that may be targeted by medulloblastoma therapies aimed at eliminating medulloblastoma recurrence.
Figures
References
-
- Baldwin C, Garnis C, Zhang L, Rosin MP, Lam WL. Multiple microalterations detected at high frequency in oral cancer. Cancer Res. 2005;65:7561–7567. - PubMed
-
- Berman DM, Karhadkar SS, Hallahan AR, Pritchard JI, Eberhart CG, Watkins DN, Chen JK, Cooper MK, Taipale J, Olson JM, et al. Medulloblastoma growth inhibition by hedgehog pathway blockade. Science. 2002;297:1559–1561. - PubMed
-
- Buttitta LA, Edgar BA. How size is controlled: From Hippos to Yorkies. Nat Cell Biol. 2007;9:1225–1227. - PubMed
-
- Calabrese C, Poppleton H, Kocak M, Hogg TL, Fuller C, Hamner B, Oh EY, Gaber MW, Finklestein D, Allen M, et al. A perivascular niche for brain tumor stem cells. Cancer Cell. 2007;11:69–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials