

Myoglobin strikes back
- PMID: 19953516
- PMCID: PMC2865719
- DOI: 10.1002/pro.300
Myoglobin strikes back
Abstract
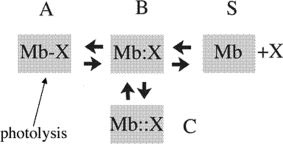
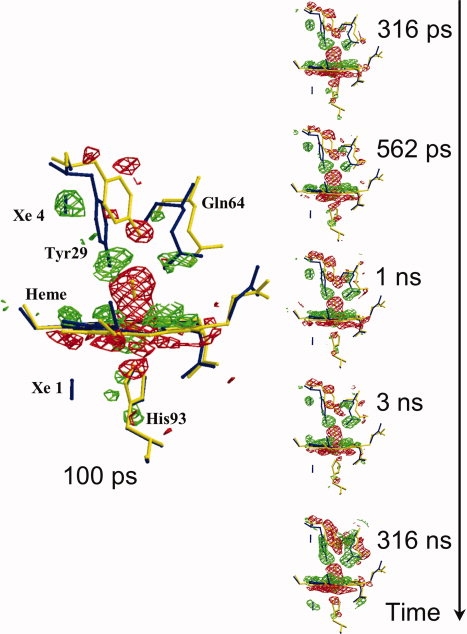
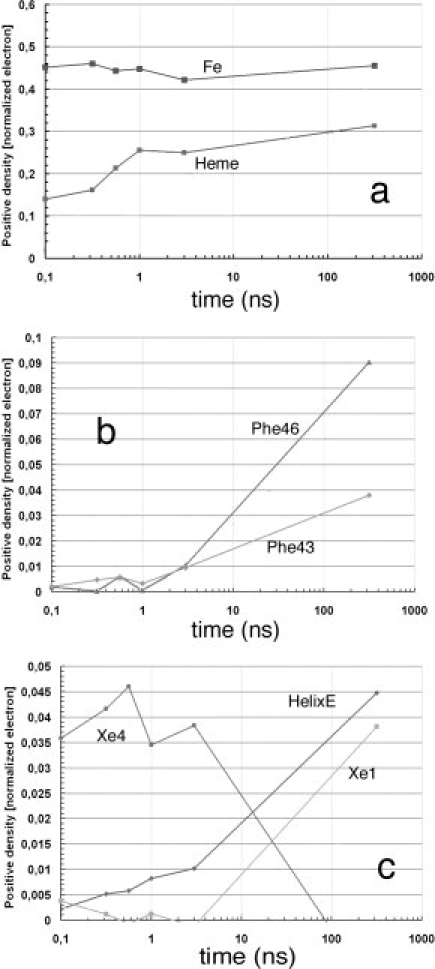
Over the last half century, myoglobin (Mb) has been an excellent model system to test a number of concepts, theories, and new experimental methods that proved valuable to investigate protein structure, function, evolution, and dynamics. Mb's function, most often considered just an oxygen repository, has considerably diversified over the last 15 years, especially because it was shown to have a role in the biochemistry of quenching and synthesizing nitric oxide in the red muscle, thereby protecting the cell. To tackle protein's structural dynamics by innovative biophysical methods, Mb has been the best prototype; laser flash technology made it possible to obtain molecular movies by time-resolved Laue crystallography (with ps resolution). This approach unveiled the complexity of the energy landscape and the structural basis of the stretched interconversion between conformational substates of a protein.
Figures
Similar articles
-
Extended subnanosecond structural dynamics of myoglobin revealed by Laue crystallography.Proc Natl Acad Sci U S A. 2006 Mar 28;103(13):4924-9. doi: 10.1073/pnas.0508880103. Epub 2006 Mar 17. Proc Natl Acad Sci U S A. 2006. PMID: 16547137 Free PMC article.
-
Photoexcitation dynamics of NO-bound ferric myoglobin investigated by femtosecond vibrational spectroscopy.J Phys Chem B. 2013 Mar 14;117(10):2850-63. doi: 10.1021/jp400055d. Epub 2013 Mar 4. J Phys Chem B. 2013. PMID: 23432208
-
Complex landscape of protein structural dynamics unveiled by nanosecond Laue crystallography.Proc Natl Acad Sci U S A. 2003 Jul 22;100(15):8704-9. doi: 10.1073/pnas.1430900100. Epub 2003 Jul 7. Proc Natl Acad Sci U S A. 2003. PMID: 12847289 Free PMC article.
-
'It's hollow': the function of pores within myoglobin.J Exp Biol. 2010 Aug 15;213(Pt 16):2748-54. doi: 10.1242/jeb.042994. J Exp Biol. 2010. PMID: 20675544 Review.
-
Recent advances in heme-protein sensors.Curr Opin Chem Biol. 2001 Apr;5(2):216-22. doi: 10.1016/s1367-5931(00)00193-9. Curr Opin Chem Biol. 2001. PMID: 11282350 Review.
Cited by
-
Dynamic multiple-scattering treatment of X-ray absorption: Parameterization of a new molecular dynamics force field for myoglobin.Struct Dyn. 2018 Sep 12;5(5):054101. doi: 10.1063/1.5031806. eCollection 2018 Sep. Struct Dyn. 2018. PMID: 30246048 Free PMC article.
-
Myoglobin in Brown Adipose Tissue: A Multifaceted Player in Thermogenesis.Cells. 2023 Sep 8;12(18):2240. doi: 10.3390/cells12182240. Cells. 2023. PMID: 37759463 Free PMC article. Review.
-
Exploring the mechanisms of the reductase activity of neuroglobin by site-directed mutagenesis of the heme distal pocket.Biochemistry. 2015 Jan 27;54(3):722-33. doi: 10.1021/bi501196k. Epub 2015 Jan 16. Biochemistry. 2015. PMID: 25554946 Free PMC article.
-
Apoprotein Intermolecular Interactions and Heme Insertion for 3D Domain Swapping in Myoglobin.ACS Omega. 2025 Feb 11;10(7):7039-7047. doi: 10.1021/acsomega.4c09948. eCollection 2025 Feb 25. ACS Omega. 2025. PMID: 40028132 Free PMC article.
-
Sulfmyoglobin Conformational Change: A Role in the Decrease of Oxy-Myoglobin Functionality.Biochem Biophys Rep. 2016 Sep;7:386-393. doi: 10.1016/j.bbrep.2016.07.002. Epub 2016 Jul 7. Biochem Biophys Rep. 2016. PMID: 28138567 Free PMC article.
References
-
- Kendrew JC, Bodo G, Dintzis HM, Parrish RG, Wyckoff HW, Phillips DC. A three-dimensional model of the myoglobin molecule obtained by X-ray analysis. Nature. 1958;181:662–666. - PubMed
-
- Bodo G, Dintzis HM, Kendrew JC, Wyckoff HW. The crystal structure of myoglobin. V. A low-resolution three-dimensional Fourier synthesis of sperm-whale myoglobin crystals. Proc R Soc Lond A. 1959;253:70–102.
-
- Kendrew JC, Dickerson RE, Strandberg BE, Hart RG, Davies DR, Phillips DC, Shore VC. Structure of myogloba three-dimensional Fourier synthesis at 2 Å resolution. Nature. 1960;185:422–427. - PubMed
-
- Millikan GA. Muscle haemoglobin. Physiol Rev. 1939;19:503–523.
-
- Wittenberg BA, Wittenberg JB. Transport of oxygen in muscle. Annu Rev Physiol. 1989;51:857–878. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
