

Toward reconstructing the evolution of advanced moths and butterflies (Lepidoptera: Ditrysia): an initial molecular study
- PMID: 19954545
- PMCID: PMC2796670
- DOI: 10.1186/1471-2148-9-280
Toward reconstructing the evolution of advanced moths and butterflies (Lepidoptera: Ditrysia): an initial molecular study
Abstract
Background: In the mega-diverse insect order Lepidoptera (butterflies and moths; 165,000 described species), deeper relationships are little understood within the clade Ditrysia, to which 98% of the species belong. To begin addressing this problem, we tested the ability of five protein-coding nuclear genes (6.7 kb total), and character subsets therein, to resolve relationships among 123 species representing 27 (of 33) superfamilies and 55 (of 100) families of Ditrysia under maximum likelihood analysis.
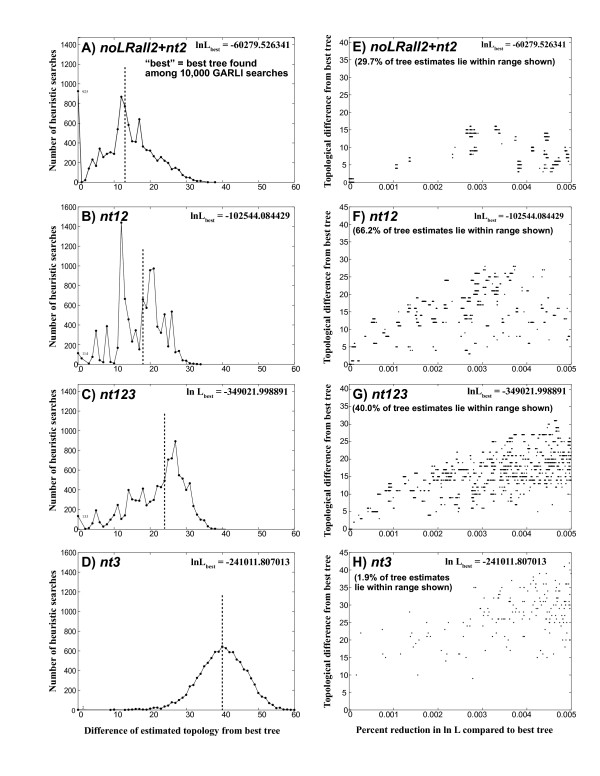
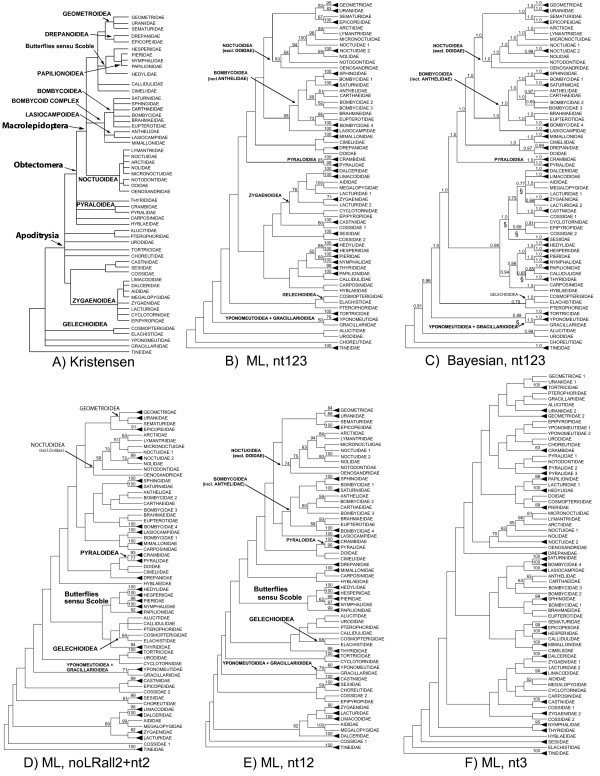
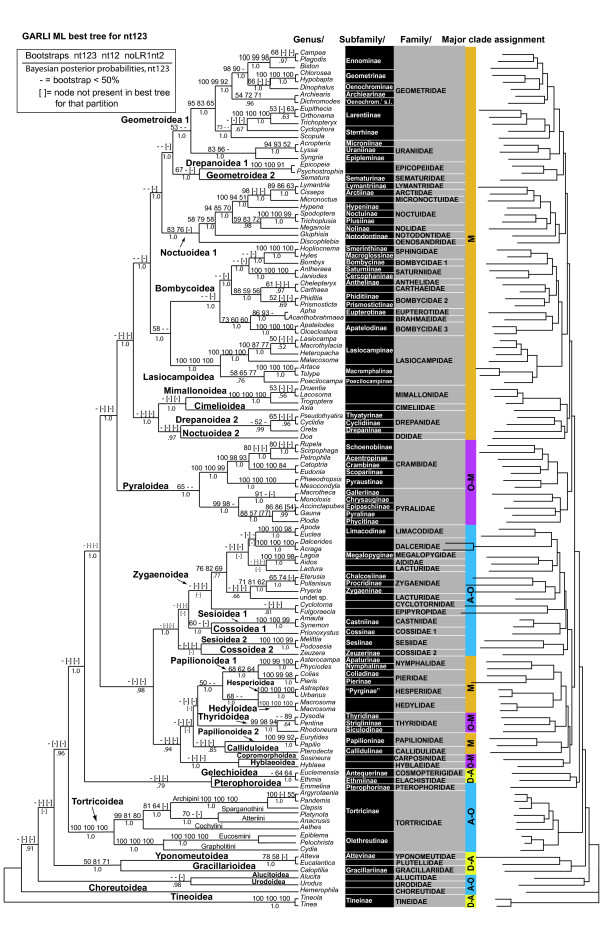
Results: Our trees show broad concordance with previous morphological hypotheses of ditrysian phylogeny, although most relationships among superfamilies are weakly supported. There are also notable surprises, such as a consistently closer relationship of Pyraloidea than of butterflies to most Macrolepidoptera. Monophyly is significantly rejected by one or more character sets for the putative clades Macrolepidoptera as currently defined (P < 0.05) and Macrolepidoptera excluding Noctuoidea and Bombycoidea sensu lato (P < or = 0.005), and nearly so for the superfamily Drepanoidea as currently defined (P < 0.08). Superfamilies are typically recovered or nearly so, but usually without strong support. Relationships within superfamilies and families, however, are often robustly resolved. We provide some of the first strong molecular evidence on deeper splits within Pyraloidea, Tortricoidea, Geometroidea, Noctuoidea and others.Separate analyses of mostly synonymous versus non-synonymous character sets revealed notable differences (though not strong conflict), including a marked influence of compositional heterogeneity on apparent signal in the third codon position (nt3). As available model partitioning methods cannot correct for this variation, we assessed overall phylogeny resolution through separate examination of trees from each character set. Exploration of "tree space" with GARLI, using grid computing, showed that hundreds of searches are typically needed to find the best-feasible phylogeny estimate for these data.
Conclusion: Our results (a) corroborate the broad outlines of the current working phylogenetic hypothesis for Ditrysia, (b) demonstrate that some prominent features of that hypothesis, including the position of the butterflies, need revision, and (c) resolve the majority of family and subfamily relationships within superfamilies as thus far sampled. Much further gene and taxon sampling will be needed, however, to strongly resolve individual deeper nodes.
Figures
References
-
- Kristensen NP, Skalski AW. In: Handbuch der Zoologie, a Natural History of the Phyla of the Animal Kingdom, Vol. IV, Arthropoda: Insecta, Part 35, Lepidoptera, Moths and Butterflies, Vol. 1: Evolution, Systematics, and Biogeography. Kristensen NP, editor. Berlin & New York: Walter de Gruyter; 1999. Phylogeny and palaeontology; pp. 7–25.
-
- Kristensen NP, Scoble M, Karsholt O. In: Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa. Zhang Z-Q, Shear WA, editor. Vol. 1668. 2007. Lepidoptera phylogeny and systematics: the state of inventorying moth and butterfly diversity; pp. 699–747.
-
- Roe AD, Weller SJ, Baixeras J, Brown J, Cummings MP, Davis D, Kawahara AY, Parr C, Regier JC, Rubinoff D, Simonsen TJ, Wahlberg N, Zwick A. In: Genetics and Molecular Biology of Lepidoptera. Goldsmith M, Marec F, editor. Boca Raton: CRC Press; 2009. Evolutionary Framework for Lepidoptera Model Systems; pp. 1–24.
-
- New TR. Invertebrate Surveys for Conservation. Oxford: Oxford University Press; 1998.
-
- Kristensen NP, (Ed) Handbuch der Zoologie, a Natural History of the Phyla of the Animal Kingdom, Vol. IV, Arthropoda: Insecta, Part 35, Lepidoptera, Moths and Butterflies, Vol. 1: Evolution, Systematics, and Biogeography. Berlin & New York: Walter de Gruyter; 1999.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
